|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
植物ESCRT复合体的功能研究进展
植物学报
2022, 57 (5):
697-712.
DOI: 10.11983/CBB22038
真核细胞中, 功能高度保守的内体蛋白分选转运装置ESCRT在胞吞途径和蛋白分泌途径中均扮演重要角色。植物细胞中, 该装置包含ESCRT-I、ESCRT-II、ESCRT-III和VPS4/SKD1复合体4个亚基, 但缺乏ESCRT-0亚基。ESCRT的每个亚基均由多个蛋白构成。目前, 针对ESCRT的研究已经证实, 其在泛素化的膜蛋白进入多囊泡体/液泡前体(MVB/PVC)内腔过程中发挥重要调控作用; 同时在自噬途径以及应对环境胁迫等方面也具有重要的调节功能。该文首先介绍了植物中ESCRT复合体的组成及生物学功能, 然后总结了植物中特有ESCRT复合体组分蛋白的最新研究进展, 最后探讨了有关ESCRT复合体研究中尚未解决的重要科学问题。
表1
内体蛋白分选转运装置(ESCRT)组分蛋白及其功能
正文中引用本图/表的段落
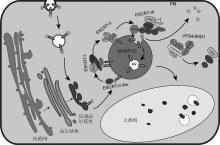
内体蛋白分选转运装置(endosomal sorting complex required for transport, ESCRT)是一个进化上高度保守且由多个亚基构成的复合体系统, 在MVB/ PVC形成及介导泛素化蛋白进入液泡降解过程中扮演重要角色(Gao et al., 2017; Isono and Kalinowska, 2017)。赵雅惠等(2017)研究发现, 在哺乳动物和酵母细胞中, ESCRT复合物均具有5个亚基, 分别为ESCRT-0、ESCRT-I、ESCRT-II、ESCRT-III和VPS4/SKD1, 每个亚基含有2-4个核心蛋白(表1)。此外, 还有多种辅助蛋白参与调节ESCRT复合物的功能。不同物种ESCRT复合物介导的泛素化蛋白降解途径可概括为以下几步: (1) ESCRT-0组分蛋白TOL识别并结合被泛素化修饰的膜蛋白; (2) 通过VPS23和VPS36的识别与结合作用, 泛素化蛋白被转移至ESCRT-I和ESCRT-II上进一步富集; (3) 通过ESCRT-II组分蛋白VPS25与ESCRT-III核心组分蛋白VPS20间的互作, ESCRT-III复合体被激活并在MVB膜上形成内陷; (4) 泛素化的膜蛋白在USP8/ Doa4作用下去泛素化后(Johnson et al., 2017)与部分MVB/PVC膜进一步内陷, 形成腔内囊泡(intraluminal vesicles, ILVs), 进入MVB/PVC内腔; (5) ESCRT-III复合体辅助蛋白招募VPS4/SKD1-LIP5复合物, 通过水解ATP将ESCRT-III复合体解聚并进入到下一个循环(图1) (夏恒传等, 2013; Gao et al., 2017; Sch?neberg et al., 2017; Schwihla and Korbei, 2020)。近年的研究发现, ESCRT复合物除介导膜蛋白降解外, 在多种其它生命活动中也发挥重要作用, 包括清理受损的核膜(Raab et al., 2016)、修复细胞膜损伤(Jimenez et al., 2014)、调控大液泡的形成(Cui et al., 2019, 2020)、参与叶绿体蛋白的选择性自噬及自噬体形成调控(Spitzer et al., 2015)和抵抗病原菌入侵等(Wang et al., 2014)。相较于哺乳动物和酵母, 模式植物拟南芥(Arabidopsis thaliana)缺少ESCRT-0复合体亚基以及ESCRT-I组分蛋白MVB12 (Gao et al., 2017), 但这并不影响ESCRT核心蛋白之间的互作网络。更重要的是, 近年研究人员鉴定到多个植物特有的ESCRT组分蛋白: FREE1 (FYVE domain protein required for endosomal sorting 1) (Gao et al., 2014, 2015)、FYVE4 (Liu et al., 2021)、PROS (positive regulator of SKD 1) (Reyes et al., 2014)、RST1 (Resurrection 1) (Zhao et al., 2015, 2019)和BRAF (BRo1-domain protein as FREE1 suppressor) (Shen et al., 2018)。这些蛋白不仅能与保守的ESCRT组分发生互作而参与调控MVB/PVC的形成, 还参与调控植物特有的生命活动, 包括植物激素信号转导通路、应答逆境胁迫以及抵御病原菌侵染等。因此, 本文首先阐述拟南芥ESCRT复合物各亚基的互作关系和功能, 之后重点讨论植物特有的ESCRT组分蛋白的功能及作用机制, 以及它们在植物细胞生命活动中的最新研究进展, 最后对植物ESCRT研究进行展望。
在动物和酵母中, ESCRT-I复合体由多个亚基按照1:1:1:1的比例组成, 包括VPS23/TSG101、VPS28、VPS37和MVB12 (MVB sorting factor of 12 kDa) (Kumar et al., 2016)。酵母中, VPS23/TSG101蛋白N端包含1个泛素结合酶E2的UEV (ubiquitin E2 variant)结构域, 该结构域在泛素化蛋白胞吞过程中发挥双重作用: 通过识别ESCRT-0亚基蛋白的P[S/T]- AP基序而与ESCRT-0相关蛋白互作, 同时识别并结合泛素化的货物蛋白。VPS28则通过其C端的四螺旋结构域与ESCRT-II组分蛋白VSP36互作, 以此连接ESCRT-I与ESCRT-II (Kostelansky et al., 2007)。研究发现, 拟南芥基因组中包含3个编码ESCRT-I亚基蛋白的基因, 其编码的亚基分别为VPS23、VPS28和VPS37。每个亚基均具有2个拷贝, 依次为VPS23A、VPS23B, VPS28A、VPS28B及VPS37-1、VPS37-2 (Schwihla and Korbei, 2020) (图3A-C)。拟南芥VPS23蛋白类似于酵母和动物中的同源蛋白, 在功能上高度保守, 既可识别并结合泛素化的货物蛋白, 也可通过与VPS28及VPS37互作形成ESCRT-I复合体。而VPS23A自身的泛素化和降解依赖能与其互作的SINAT (SEVEN IN ABSENTIA OF ARABIDOPSIS THALIANA)家族蛋白(Xia et al., 2020)。目前, 针对VPS23A功能的研究发现, VPS23A蛋白的表达水平对ABA (abscisic acid)信号通路受体蛋白PYR1 (PYRABACTIN RESISTANCE1)的亚细胞定位和PYL4 (PYR1-LIKE 4)的稳定性均具有重要调控作用(Yu et al., 2016)。因此, VPS23A通过调节PYR1/ PYL4蛋白的分选与降解进而负调控ABA信号通路。更重要的是, VPS23A不仅与ESCRT-III复合体相关蛋白ALIX (ALG-2 INTERACTING PROTEIN-X)互作, 也可结合植物特有的ESCRT组分蛋白FREE1。对ALIX的研究发现, 其具有促进ABA信号受体蛋白PYR/PYL/RCAR (REGULATORY COMPONENTS OF ABA RECEPTORS)降解和控制气孔开合角度的作用, ALIX基因缺失会导致VPS23A结合ABA信号受体蛋白的能力显著降低(García-León et al., 2019)。Gao等(2014)和Belda-Palazon等(2016)对FREE1的研究表明, FREE1也参与调控ABA信号受体蛋白PYL4进入液泡的降解过程, 可见ESCRT-I及ESCRT相关蛋白对植物激素信号通路具有调节作用。进一步研究发现, VPS23A还参与植物的抗逆反应, 其通过调控SOS2 (Salt-Overly-Sensitive2)的质膜定位及SOS2与SOS3间的结合强度参与植物抵抗盐胁迫的应答过程(Yu et al., 2016)。近期, Liu等(2020)针对VPS28的研究发现, 其亚细胞定位于TGN和MVB/ PVC并在胚胎形成期高水平表达。VPS28A和VPS28B蛋白在功能上存在冗余, vps28a或vps28b突变体无明显的表型缺陷, 但是vps28a/vps28b双突变体的胚对生长素表现不敏感, 胚的发育在球型胚阶段被抑制。进一步观察发现, 双突变体的液泡形态异常, 生长素转运载体PIN1 (PIN-FORMED 1)异常积累在液泡膜(Liu et al., 2020), 表明VPS28对液泡发育及膜蛋白的内吞具有重要作用。在植物抵御病原菌侵染时, vps28b以及vps37-1突变体中flg22 (flagellin 22)诱导的受体激酶FLS2 (flagellin sensing 2)的内吞过程受到显著抑制, 突变体表现出感病性(Spallek et al., 2013), 表明ESCRT-I不仅参与调控胚胎发育、 MVB/PVC的形成和膜蛋白的内吞, 也参与调控植物激素信号通路的开关和应对细菌病原体的免疫应答过程。
ESCRT-II作为ESCRT复合物行使功能的分子枢纽, 在功能上起着承上启下的作用。ESCRT-II复合体是分子量为155 kDa的异四聚体, 由VPS22/EAP30、VPS36/EAP45和2个VPS25/EAP20亚基组成, 在进化过程中高度保守(Morita et al., 2007; Spallek et al., 2013), 不同物种中均已鉴定出对应的同源蛋白。对酵母ESCRT-II的结构分析发现, ESCRT-II复合体呈现“Y”型结构, 该结构包含1个分子的VPS22, 2个分子的VPS25和长度为171 aa的VPS36碳端结构域。在此结构中, VPS22蛋白序列中的CC (coiled-coil)结构域通过结合VPS36蛋白的C端结构域形成“Y”型结构的1个分支, 同时VPS36和VPS22分别结合1个VPS25分子形成“Y”型结构的另一个分支和主轴(Hierro et al., 2004)。通过对ESCRT-II组分蛋白的功能进行研究, 发现VPS25通过与ESCRT-III复合体中的VPS20互作进而调节ESCRT-III复合体组分的招募和复合体的形成(Babst et al., 2002)。VPS36则通过其N端结构域与ESCRT-I复合体中的VPS28互作而识别泛素化蛋白。对拟南芥VPS25的研究发现, VPS25的表达水平受生长素诱导, vps25突变体表现出对生长素敏感, 用浓度为0.1 mg·L-1生长素处理, 突变体主根的生长受到强烈抑制(Guo et al., 2014); 对VPS36的研究表明, 其基因缺失突变会引起胚发育迟缓、根生长缓慢及子叶表皮细胞形态异常等表型。突变体细胞中晚期内吞体异常积累, MVB/PVC和液泡的形成均受到抑制, 生长素转运载体PIN1、PIN2以及AUX1 (AUXIN RESISTANT 1)的内吞过程与定位均受到严重影响(Wang et al., 2017)。敲除水稻(Oryza sativa) OsVPS22基因则会导致主根生长受到严重抑制, 胚乳无法正常灌浆, 最终突变体幼苗无法存活(Zhang et al., 2013), 因此ESCRT-II组分蛋白在植物生长发育过程中发挥重要作用。
在酵母和动物细胞中, ESCRT-III由4个亚基组成, 即VPS2p/CHMP2 (charged multivesicular body protein 2)、VPS20p/CHMP6、VPS24p/CHMP3和SNF7p (sucrose non-fermenting 7)/CHMP4。此外, 还包括Did2 (Doa4-independent degradation2)/CHMP1、VPS60/CHMP5和IST1 (increased salt tolerance 1) 3个辅助蛋白(Teis et al., 2008)。在内吞途径中, ESCRT-III组分蛋白VPS20首先与ESCRT-II组分蛋白VPS25互作而激活ESCRT-III, 同时SNF7多聚体被VPS20招募到MVB/PVC膜上组装ESCRT-III, 并进一步招募VPS2和VPS24等蛋白到MVB/PVC膜形成成熟的ESCRT-III复合体, 从而介导膜的凹陷和剪切形成ILVs。Did2/CHMP1、VPS60/CHMP5和IST1三个辅助蛋白则通过激活VPS4/SKD1-LIP5复合体活性参与调节ESCRT-III与VPS4/SKD1-LIP5复合体结合的强弱、ESCRT-III从MVB/PVC膜上的解离及解聚过程, 以此调控ESCRT-III组分蛋白的循环利用和MVB/PVC的成熟(Henne et al., 2011)。拟南芥中的ESCRT-III具有高度保守性, 对应的同源蛋白均含有多个亚基, 包括VPS2.1、VPS2.2和VPS2.3, VPS20.1和VPS20.2, SNF7A和SNF7B, VPS24-1和VPS24-2 (Gao et al., 2017)以及能够与SNF7发生互作的ALIX (图4A-D)。通过酵母双杂交筛选, 发现ESCRT-III组分蛋白VPS2.1和VPS24.1均可与去泛素化酶AMSH3 (associated molecular with SH3 domain of STAM 3)互作(Katsiarimpa et al., 2011), 它们之间的互作分别依赖AMSH3的MIT (microtubule interacting and transport)结构域和ESCRT-III的MIM1 (microtubule interacting and transport (MIT)-interacting motif1)结构域。亚细胞定位研究显示, AMSH3与ESCRT-III组分蛋白共定位于E类晚期胞内吞小体(class E compartments)中, AMSH3的DUB (deubiquitinating enzyme)结构域缺失会导致ESCRT-III亚细胞定位错乱, 因此AMSH3对ESCRT-III正确的亚细胞定位具有重要作用(Katsiarimpa et al., 2011)。AMSH3定位到晚期内吞体则受到其互作蛋白ALIX的调控, ALIX基因缺失突变不仅会导致植株胚死亡, MVB/PVC和液泡形态异常, 而且会使AMSH3定位到晚期内吞体的能力消失(Kalinowska et al., 2015)。由此可见, AMSH3与ESCRT-III的协同作用依赖ALIX的功能。对AMSH3基因功能的进一步研究显示, 缺失AMSH3基因也会引起类似alix突变体的表型, 并进一步表现出泛素化蛋白和自噬体异常积累等表型(Isono et al., 2010)。此外, 对vps2.1突变体的研究发现, 突变体植株表现出胚死亡, 具有类似于chmp1a/chmp1b双突变体的表型(Spitzer et al., 2009; Katsiarimpa et al., 2011), 因此ESCRT-III复合体组分蛋白的缺失均可导致植物发育受阻而死亡。近期研究发现, 另一个去泛素化酶AMSH1也可识别并结合VPS2.1, 参与调节货物蛋白的去泛素化过程。过表达VPS2.1或缺失AMSH1基因均会导致植物出现矮化和早衰表型, 并表现出PIN2异常积累于液泡膜及其内吞降解过程受到强烈抑制, 自噬体标记物ATG8e异常积累以及自噬途径受阻等现象(Katsiarimpa et al., 2011, 2013; Schwihla and Korbei, 2020), 表明VPS2.1和2个去泛素化酶AMSH3与AMSH1在植物生长及物质循环利用过程中起重要作用。在拟南芥悬浮细胞中, 过表达活性缺失形式的vps24.1-1-152会导致MVB/PVC内腔中ILV数量显著减少, MVB/PVC形态异常(Cai et al., 2014), 同时导致生长素生物合成关键酶YUC1黄素单氧合酶(YUC1 flavin monooxygenase)无法正常进入液泡降解(Ge et al., 2019)。进一步研究发现, ESCRT-III辅助蛋白CHMP1可与VPS4/SKD1结合, 调节MVB/ PVC的形成及ESCRT-III复合体的解聚。chmp1a/ chmp1b双突变体植株中, 生长素转运载体蛋白PIN1、PIN2和AUX1在细胞膜上积累, 无法正常内吞降解, 依赖于自噬途径降解的质体(plastid)蛋白大量积累于细胞质; 并表现出胚胎或早期幼苗致死表型(Spitzer et al., 2009, 2015)。因此, ESCRT-III组分蛋白对植物是否能够存活具有重要作用; 同时, ESCRT- III组分蛋白的缺失会导致自噬体积累, 进而使依赖于自噬体途径降解的蛋白异常积累。但截至目前, 人们对ESCRT-III和自噬体协同作用的调控因子仍知之甚少, 有待进一步探究。
VPS4/SKD1-LIP5具有ATPase活性, 通过水解ATP使ESCRT-III复合体从MVB/PVC膜上脱落, 解离进入下一个循环, 同时促进ILV囊泡化。在此过程中, LIP5 (LYST-INTERACTING PROTEIN5)可与VPS4/ SKD1互作形成复合体; 而体外ATP水解酶活性实验显示, LIP5可将SKD1对ATP的水解能力提高4-5倍。因此, LIP5作为SKD1活性的重要调控因子可通过正向调控VPS4/SKD1ATPase活性及VPS4/SKD1低聚结构的稳定性而参与对ESCRT-III复合体解聚的调控(Haas et al., 2007; Norgan et al., 2013)。在烟草(Nicotiana tabacum)悬浮细胞BY-2 (Bright Yellow 2)中, 过表达SKD1 ATPase活性缺失突变形式的SKD1E232Q不仅造成MVB形态异常, 还会导致细胞坏死; 在拟南芥中过表达SKD1E232Q则会引起MVB膨大和腔内ILV数量显著减少等表型(Haas et al., 2007)。进一步研究显示, 过表达SKD1E232Q会导致ESCRT- III组分蛋白VPS2.1以及ALIX积累在膨大的囊泡(enlarged vesicular compartments)无法正常行使功能(Kalinowska et al., 2015)。因此, 精准调控VPS4/ SKD1的表达水平对确保ESCRT正常行使功能具有重要意义。通过对skd1以及lip5突变体的研究发现, SKD1基因缺失会导致植株死亡, 而lip5突变体植株与野生型无明显差异(Haas et al., 2007; Buono et al., 2016), 但在lip5突变体中PIN2和PIN3均异常积累于液泡膜(Buono et al., 2016)。Xia等(2016)研究发现, 过表达LIP5会增强植株对ABA的超敏反应以及对干旱的抵抗能力, 同时促进气孔关闭。而当受到病原菌PstDC3000 (Pseudomonas syringae pv. to-mato strain DC3000)侵染时, LIP5可以被蛋白激酶MPK3 (mitogen-activated protein kinases 3)以及MPK6磷酸化, 正向调控MVB/PVC在病原菌侵染点快速形成以抵御病原菌的入侵(Wang et al., 2014)。因此, VPS4/SKD1-LIP5复合体在植物生长发育、抵御外界不利环境和病原菌免疫应答过程中均发挥重要作用。但是, VPS4/SKD1-LIP5复合体参与ABA途径的具体机制目前尚不清楚, 有待深入研究。
由于植物中缺乏类似于动物和酵母的ESCRT-0组分同源蛋白, 因此有无蛋白代替ESCRT-0复合体识别、结合泛素化的货物蛋白并招募下游ESCRT组分蛋白, 成为研究植物ESCRT复合物的重要问题之一(崔勇等, 2019)。通过对ESCRT-0复合体亚基VPS27/HRS保守结构域氨基酸序列的比对分析, 在拟南芥中鉴定到含有该保守结构域的FREE1蛋白(图2)。FREE1是双子叶植物中的特有蛋白, 在酵母和动物中均未发现其同源蛋白。FREE1蛋白由N端的Pro富集结构域(PRR)、C端的FYVE结构域和CC结构域组成, 在FYVE结构域中还存在2个PTAP-like基序。FYVE结构域在功能上高度保守, 通过PTAP-like基序, FREE1能与ESCRT-I组分蛋白VPS23互作, 也可以与MVB/ PVC膜上的磷脂酰肌醇3-磷酸(phosphatidylinositol- 3-phosphate, PI3P)结合从而定位到MVB/PVC膜上。此外, 通过其N端的PRR结构域, FREE1蛋白也可以与在ESCRT-I上发挥功能的ALIX蛋白互作, 调控MVB/PVC的形成(Shen et al., 2016)。FREE1基因缺失会导致MVB/PVC无法正常形成, 内腔中ILV数量显著减少, 大量小液泡堆积无法形成正常形态以及功能的大液泡, 致使泛素化蛋白降解受阻, 自噬体大量积累, 最终导致植株死亡。因此, FREE1蛋白对ESCRT介导的泛素化蛋白质降解起重要调控作用(Gao et al., 2014, 2015)。此外, FREE1蛋白还可通过其碳端的CC结构域与自噬途径调节因子SH3P2 (SH3 domain-containing protein 2)蛋白氮端的BAR (Bin- amphiphysin-Rvs)结构域相互作用, 介导自噬体形成, 调节蛋白质的降解(Gao et al., 2015)。Belda- Palazon等(2016)针对FREE1功能的进一步研究发现, FREE1不仅能介导ABA受体蛋白PYL4进入液泡降解, 在ABA处理条件下, FREE1蛋白还可被SnRK2 (SNF1-related protein kinase 2)蛋白激酶磷酸化, 进入细胞核与碱性亮氨酸拉链转录因子ABI5 (ABA- INSENSITIVE5)和ABF4 (ABA-RESPONSIVE ELEMENTS BINDING FACTOR 4)结合, 进而抑制其活性, 降低其结合下游基因的能力, 抑制下游基因的表达水平。利用CRISPR-Cas9技术获得的free1-ctmut突变体则表现出ABA敏感性(Li et al., 2019), 暗示FREE1除在传统囊泡运输过程及自噬体途径发挥作用, 还在ESCRT介导的胁迫应激反应中行使重要功能。Xia等(2020)和Xiao等(2020)研究表明, FREE1与SINAT家族蛋白SINAT1-5均具有互作关系, SINAT可通过调控FREE1及ESCRT-I组分蛋白VPS23A的泛素化而进入蛋白酶体降解途径。不仅如此, SINAT还可结合FREE1或VPS23A, 形成SINAT-FREE1或SINAT-VPS23A复合体, 该复合体的降解依赖MVB/ PVC介导的自噬体途径(Xia et al., 2020)。因此, SINAT蛋白可能是决定ESCRT组分蛋白进入蛋白酶体或自噬体降解途径的关键因子。Xiao等(2020)进一步研究显示, 过表达SINAT1-4蛋白可促进FREE1降解, 但过表达SINAT5可增强FREE1的稳定性, 因此SINAT家族蛋白在调控ESCRT-I复合体功能上具有多样性和精密性。
植物特有的ESCRT复合物组分蛋白FREE1在多囊泡体形成和膜蛋白进入液泡降解途径中发挥重要作用。为更深入了解FREE1蛋白在植物生长发育等生物学过程中的功能, Zhao等(2015)利用EMS (ethyl met- hane sulfonate)对free1 RNAi突变体进行诱变处理, 通过表型分析筛选出FREE1功能恢复突变体, 即sof (suppressor of free1)。后续通过全基因组测序分析和细胞内液泡形态染色观察等一系列实验手段, 鉴定出多个潜在的引起sof突变的基因, 包括BRAF和RST1 (Zhao et al., 2015)。
根据氨基酸序列分析, 含有401个氨基酸的BRAF蛋白属于具有BRO1结构域的蛋白家族, 其N端存在S-酰化修饰位点。对BRAF亚细胞定位的研究显示, BRAF依赖的S-酰化修饰位点主要定位于细胞膜, 但通过与ESCRT-I组分蛋白FREE1和VPS23互作, 一部分BRAF也与FREE1和VPS23共定位于MVB/PVC膜上。进一步研究发现, BRAF和FREE1可竞争性结合VPS23蛋白的CC结构域, 过表达BRAF会导致FREE1结合MVB/PVC膜的能力显著降低。据此, Shen等(2018)阐述了BRAF负调控FREE1的功能进而参与调控MVB/PVC形成的分子机理。在植物逆境胁迫应答中, 大量依赖S-酰化修饰而定位在细胞膜上的蛋白会从膜上脱落, 进而诱导逆境胁迫应答相关基因的表达, 如调控植物应对病原菌侵染的RIN4 (RPM1-interacting protein 4)蛋白。RIN4通过S-酰化修饰定位于细胞膜, 受到病原菌侵染时, RIN4被磷酸化, 从细胞膜脱离进入细胞质, 最终激活RPM1介导的免疫反应(Mackey et al., 2002)。BRAF蛋白也具有类似RIN4的生理特点, 如N端具有S-酰化修饰及S-酰化位点突变导致植物死亡。因此, 推测BRAF可能在某些环境或生长条件下, 通过S-酰化过程调节BRAF蛋白在细胞质、细胞膜和多囊泡体中的分布, 以应对环境胁迫, 但目前针对BRAF调控逆境胁迫反应的分子机理还有待深入研究(Shen et al., 2018)。
与BRAF具有负调控FREE1的特点类似, RST1基因突变也能恢复free1 RNAi突变体的致死表型, 因此RST1是FREE1功能的抑制因子之一。RST1蛋白包含1 841个氨基酸, 存在于多个物种中。对RST1的研究显示, 该蛋白与拟南芥茎角质层蜡的分布、胚胎发育和对病原菌的免疫反应有关(Chen et al., 2005; Mang et al., 2009)。而通过EMS诱变, 在free1 RNAi突变体背景下, RST1基因功能缺失可恢复free1 RNAi突变体植株的致死表型, MVB/PVC和液泡均能正常发育且具有正常的生物学功能(Zhao et al., 2015)。对RST1蛋白的亚细胞定位研究发现, 不同于传统的ESCRT组分蛋白定位于MVB/PVC, RST1完全定位于细胞质中, 与FREE1蛋白无明显的共定位关系。过表达RST1会导致植株表现出生长受抑制、MVB/PVC异常增大及货物蛋白到液泡的运输能力减弱等表型, 表明RST1通过负调控FREE1蛋白参与ESCRT介导的MVB/PVC形成途径(Zhao et al., 2019)。Zhao等(2019)进一步对free1 RNAi和rst1突变体转录组RNA-seq分析显示, TOL4蛋白表达水平在2个突变体植株中存在显著差异, 在free1 RNAi中表达水平异常升高, 而在rst1中却显著降低, 暗示TOL4蛋白可能与FREE1和RST1协同作用于MVB/PVC的形成和液泡转运过程, 但具体作用机理尚需进一步探索。
本文的其它图/表
|
 首页
首页