|
|
||||||||||||||||
|
沙棘hrh-miRn458靶向转录因子WRI1调控油脂合成
植物学报
2022, 57 (5):
635-648.
DOI: 10.11983/CBB22004
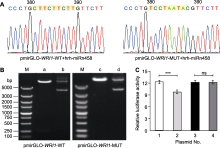
转录因子WRI1 (WRINKLED 1)在油脂合成过程中发挥重要调控作用, 其转录和翻译水平调控机理以及下游的靶基因现已基本明确, 但尚未见其转录后调控的报道。为探讨沙棘(Hippophae rhamnoides) hrh-miRn458与转录因子WRI1之间的靶向关系, 并阐明其在果肉和种子发育过程中的表达模式, 通过生物信息学方法预测hrh-miRn458成熟体序列及其与候选靶基因WRI1的结合位点, 采用双荧光素酶报告基因检测和RNA pull down技术相结合的方法验证hrh-miRn458与WRI1基因之间的靶向关系, 并应用qRT-PCR方法分析hrh-miRn458与WRI1在沙棘不同发育期果肉和种子油脂合成过程中的表达变化。结果表明, 生物信息学预测WRI1基因的CDS区第309-327位与hrh-miRn458成熟体序列的15个碱基互补; 荧光检测显示pmirGLO-WRI1-WT+hrh-miRn458 mimics显著抑制荧光素酶活性(P<0.001); RNA pull down实验证实hrh-miRn458与WRI1存在互作关系; 不同发育期果肉和种子中hrh-miRn458的表达量总体呈先下降后上升趋势, 其靶基因WRI1的表达量则呈先上升后降低趋势; 同一时期, 果肉中hrh-miRn458的表达量明显低于种子, 而果肉中的WRI1表达量则明显高于种子。综上, 沙棘hrh-miRn458靶向转录因子WRI1, 且二者在果肉和种子油脂合成过程中存在负调控关系。研究结果为深入理解沙棘种子油脂合成机制及培育高油品种提供了参考依据。
表1
qRT-PCR引物
正文中引用本图/表的段落
摘要:转录因子WRI1 (WRINKLED 1)在油脂合成过程中发挥重要调控作用, 其转录和翻译水平调控机理以及下游的靶基因现已基本明确, 但尚未见其转录后调控的报道。为探讨沙棘(Hippophae rhamnoides) hrh-miRn458与转录因子WRI1之间的靶向关系, 并阐明其在果肉和种子发育过程中的表达模式, 通过生物信息学方法预测hrh-miRn458成熟体序列及其与候选靶基因WRI1的结合位点, 采用双荧光素酶报告基因检测和RNA pull down技术相结合的方法验证hrh-miRn458与WRI1基因之间的靶向关系, 并应用qRT-PCR方法分析hrh-miRn458与WRI1在沙棘不同发育期果肉和种子油脂合成过程中的表达变化。结果表明, 生物信息学预测WRI1基因的CDS区第309-327位与hrh-miRn458成熟体序列的15个碱基互补; 荧光检测显示pmirGLO-WRI1-WT+hrh-miRn458 mimics显著抑制荧光素酶活性(P<0.001); RNA pull down实验证实hrh-miRn458与WRI1存在互作关系; 不同发育期果肉和种子中hrh-miRn458的表达量总体呈先下降后上升趋势, 其靶基因WRI1的表达量则呈先上升后降低趋势; 同一时期, 果肉中hrh-miRn458的表达量明显低于种子, 而果肉中的WRI1表达量则明显高于种子。综上, 沙棘hrh-miRn458靶向转录因子WRI1, 且二者在果肉和种子油脂合成过程中存在负调控关系。研究结果为深入理解沙棘种子油脂合成机制及培育高油品种提供了参考依据。
WRI1属于植物特有的AP2类转录因子, 被认为是提高油脂产量的最有效转录因子(Kong et al., 2019; Rich et al., 2021)。自1998年转录因子WRI1首次在拟南芥褶皱突变体中被发现以来, 围绕WRI1在油脂合成中的生物学功能展开了大量研究, 基本明确了转录因子WRI1上游的转录调控、翻译后水平的修饰以及下游靶基因等一系列调控机制(金龙飞等, 2022)。过表达WRI1显著提高转基因植株的种子含油率。例如, 在拟南芥wri1突变体中过表达亚麻荠(Camelina sativa) WRI1恢复了突变体的种子表型且油脂含量增加2.5-4.0倍(An et al., 2017); 过表达蓖麻(Ricinus communis) RcWRI1的烟草(Nicotiana tabacum)叶片含油率升高了4.3-4.9倍(Ji et al., 2018)。目前已明确WRI1的上游调控转录因子主要包括正调控因子ABI3、FUS3、LEC1、LEC2和NY-YA3 (NUCLEAR FACTOR Y submit A3)及负调控因子MYB89和TCP等(Kong et al., 2019)。例如, 油棕榈的EgNF-YA3与EgWRI1-1直接互作, 激活下游脂肪酸合成基因的表达, 促进油脂积累(Yeap et al., 2017)。在翻译后水平的微调控方面, 目前已发现与WRI1互作的蛋白主要包括正调控蛋白14-3-3、MED15 (Mediator 15)和APUM24 (PUMILIO PROTEIN24)及负调控蛋白BPM1 (BTB/POZMATH 1)、KIN10激酶和TCP4等(Kong et al., 2019)。例如, 超表达14-3-3显著提高拟南芥转基因植株AtWRI1的表达水平(Ma et al., 2016); KIN10介导AtWRI1的70位点苏氨酸和166位点丝氨酸的磷酸化降解, 进而负调控油脂合成(Zhai et al., 2017)。此外, 通过分析过表达和突变体中的差异表达基因, 预测转录因子WRI1下游涉及糖酵解、脂肪酸合成及调控TAG (triacylglyceride)组装和油体贮存的相关靶基因。过表达JcWRI1激活了麻枫树(Jatropha curcas)种子中PDHα (pyruvate dehydrogenase)、PKpα (pyruvate kinase)、PKpβ、BCCP2 (biotin carboxyl carrier protein 2)、KASI、KASIII、FATA (stearoyl-acyl carrier protein thioesterase A)、FATB1、FATB2、ACP1 (Acyl carrier protein 1)和DGAT1基因的上调表达(Ye et al., 2018)。但截至目前, 未见非编码RNA直接参与转录因子WRI1转录后调控的报道(Cheng et al., 2021)。我们前期对沙棘不同发育期种子miRNA-seq和mRNA- seq数据进行联合分析, 发现沙棘hrh-miRn458靶向转录因子WRI1, 但并未验证沙棘hrh-miRn458和转录因子WRI1之间的靶向关系, 也未分析二者在不同发育期沙棘果肉和种子中的表达情况(Li et al., 2019)。
提取沙棘杂56不同发育期种子和果肉的总RNA (参照柱式植物总RNA抽提纯化试剂盒(上海生工)说明书), 合成cDNA第1链(参照Mix-X miRNA First-Stand Symthesis Kit试剂盒(大连宝生生物)说明书)。qRT- PCR分析所需的特异引物通过Primer Premier 5在线软件设计(表1), 其中UBQ5为目的基因WRI1表达水平分析的内参基因, U6为hrh-miRn458表达水平分析的内参基因。采用Mir-X miRNA qRT-PCR TB Green Kit试剂盒(大连宝生生物)和ABI-7500 RT-PCR仪(Applied Biosystems, 美国)推荐方法进行qRT-PCR实验。PCR反应体系总体积为25 μL: 12.5 μL TB Green Advantage Premix (2×), 2 μL cDNA, 正、反向引物(10 μmol·L-1)各0.5 μL, 0.5 μL ROX Dye (50×)以及9 μL RNase Free ddH2O。PCR反应程序: 95°C预变性10秒; 95°C5秒, 60°C20秒, 40个循环。溶解曲线: 95°C60秒, 55°C1分钟, 95°C30秒。用2-ΔΔCT方法分析目的基因或miRNA的相对表达量, 实验设3次重复。用SPSS 26.0软件进行单因素方差分析, 用LSD法进行差异显著性检验(P<0.05), 用Origin 8.5软件作图。
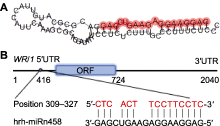
沙棘hrh-miRn458前体序列长度为74 nt, 可以形成稳定的茎环结构(图2A), 成熟体序列(5′-GAGGAA- GGAGAAGUCGAG-3′)产生于前体茎环结构5′端臂上, 最小自由能为-23.6 kcal·moL-1。经psRNATarget软件预测, 发现WRI1基因CDS区第309-327位与hrh-miRn458成熟体序列的15个碱基互补, 表明沙棘WRI1可能是hrh-miRn458的潜在靶基因(图2B)。
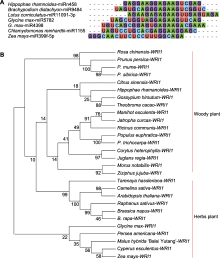
Hrh-miRn458为沙棘所特有, 其与植物中序列相似性相对较高的二穗短柄草(Brachypodium distachyon) bdi-miR9484和百脉根(Lotus corniculatus) lja-miR11091-3p的成熟体序列间仅有13个碱基相同, 与大豆的gma-miR5782和gma-miR4398成熟体序列间仅有12个碱基相同(图3A)。沙棘HrWAR1与其它27个草本和木本植物的WRI1同源序列间的系统进化关系分析表明, 沙棘HrWRI1与可可豆(Theobroma cacao)的TcWRI1及陆地棉(Gossypium hirsutum)的GhWRI1亲缘关系较近, 最先聚为一支, 然后与柑橘(Citrus sinensis)的CsWRI1聚在一起, 而与大豆的GmWRI1、鳄梨(Persea americana)的PaWRI1、芭蕾玉棠苹果(Malus hybrida ‘Balei Yutang’)的McWRI1、油莎草(Cyperus esculentus)的CeWRI1和玉米(Zea mays)的ZmWRI1的亲缘关系则相对较远(图3B)。
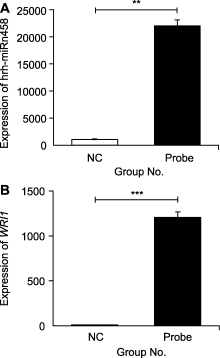
探针富集hrh-miRn458效果的qRT-PCR检测结果表明, NC组和探针组均富集到hrh-miRn458, 探针组中hrh-miRn458的表达量显著高于NC组(图5A), 说明磁珠通过生物素biotin探针拉下了体系中的hrh- miRn458。hrh-miRn458与WRI1间互作关系的qRT- PCR检测结果表明, 探针组中WRl1的表达量显著高于NC组(图5B), 说明hrh-miRn458拉下了体系中的WRl1, hrh-miRn458与WRl1之间存在互作关系。
(A) 探针组和NC组富集hrh-miRn458的表达水平; (B) 探针组和NC组富集WRI1的表达水平。 **和***分别表示NC组和探针组富集hrh-miRn458与WRI1间表达在0.01和0.001水平上差异显著。
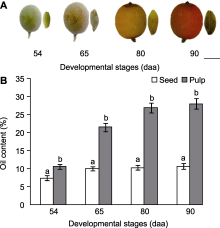
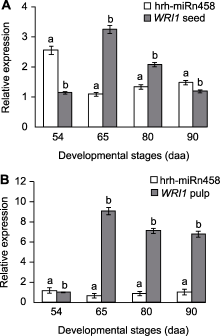
在沙棘果实的种子和非种子组织(果肉)中, 花后54天, hrh-miRn458在种子中的表达量是果肉中的2.29倍; 花后65天, hrh-miRn458在种子中的表达量是果肉中的1.80倍, 而同一时期其靶基因WRI1在种子中的表达量则比果肉中下降了64.13%; 花后80和90天, hrh-miRn458在种子中的表达量分别是果肉的1.66倍和1.52倍, 而其靶基因WRI1在种子中的表达量则分别比果肉中下降了70.56%和82.41%。
对比分析hrh-miRn458及其靶基因WRI1在果肉和种子中的表达量变化, 表明沙棘hrh-miRn458负调控其靶基因WRI1, 当果肉和种子中hrh-miRn458的表达量达到不同发育期的最高水平时, 其靶基因WRI1的表达量则为不同发育期的最低值。
自从Focks和Benning (1998)的研究发现wri1突变体的油脂含量降低80%以来, 对油脂合成主调节器之转录因子WRI1的研究一直是油脂合成机制解析的热点。油棕(Phoenix dactylifera)的EgWRI1-1在果肉中高表达, 而EgWRI1-2和EgWRI1-3则在种子中高表达(Yeap et al., 2017)。乌桕(Triadica sebifera) TsWRI1-1基因主要在种子中表达, 而TsWRI1-2基因则主要在果肉中表达(Divi et al., 2016)。本研究发现, 沙棘不同发育期果肉和种子中WRI1的表达量存在明显差异, 且在高含油率果肉中的表达量显著高于低含油率的种子。这与前期报道的WRI1表达量与种子含油率密切相关的研究结果一致。含油率高的蓖麻和燕麦(Avena sativa)的WRI1表达量明显高于淀粉含量高的小麦(Triticum aestivum) (Yang et al., 2019); WRI1在高含油率的黄油莎草(Cyperus esculentus)中的表达量为低含油率紫油莎草(C. rotundus)的14.3倍(Ji et al., 2021); 高含油率油茶品种中WRI1的表达量也明显高于低含油率的油茶品种(Wu et al., 2019)。同时, 沙棘的WRI1与可可豆(含油53%- 58%)、麻风树(种仁含油60%)、榛子(种仁含油60.5%)和核桃(含油65%-70%)等含油率高的木本油料树种的同源WRI1序列聚类在一个分支上, 暗示了通过转录因子WRI1改良沙棘种子含油率的潜力, 但其实际效果和调控机制仍有待深入研究。
miRNA介导的转录后调控在油脂合成中发挥重要作用。大豆miRNA159和miRNA319通过抑制MYB33表达下调淀粉合成相关基因APL (glucose- 1-phosphate adenylyltransferase)、AMY2 (amylase 2)、ISA2 (isoamylase 2)、ISA3、SUC (sucrose transporters)和SUS (sucrose synthase)的表达, 从而促进更多的碳代谢流向油脂合成(Cheng et al., 2021)。油棕榈miRNA5179通过抑制EgMADS16表达上调EgMADS16下游靶基因EgFAD2、EgSAD和EgDGAT2的表达, 从而促进硬脂酸和亚麻酸合成及油脂积累(Wang et al., 2021b)。沉默miR319提高TCP4的表达量, 负调控AtWRI1的表达, 降低了种子含油率(Kong et al., 2020)。本研究通过miRNA保守性分析表明, hrh-miRn458为沙棘所特有, 不仅在拟南芥中未发现与沙棘hrh-miRn458成熟体相近的miRNA, 而且它与同源性相对较高的二穗短柄草bdi- miR9484和百脉根lja-miR11091-3p的成熟体序列间也仅有13个碱基相同。同时, 沙棘hrh-miRn458靶向WRI1的CDS区, 这与植物miRNA主要结合靶基因CDS区的报道相一致(张翠桔等, 2020)。结合不同发育期果肉和种子中hrh-miRn458和WRI1表达量分析, 表明二者存在负调控关系, 这与诸多关于靶向CDS区的miRNA调控靶基因表达的研究结果相一致(Zhang et al., 2018b)。例如, bna-miR156靶向BnaSBP (SQUAMOSA promoter-binding protein)编码基因5′UTR调控油菜生长发育过程(李晓康等, 2018); 毛竹(Phyllostachys edulis)的miR164b对靶点位于编码区的靶基因PeNAC1具有表达调控作用(王丽丽等, 2015); 靶位点位于CDS区的ssa-mir-196a-4负调控靶基因lgr8 (leucine-rich G protein-coupled receptor 8)的表达(Tang et al., 2022)。Cheng等(2021)对大豆639个miRNA与WRI1之间的靶向关系进行了预测, 但未发现靶向WRI1的miRNA; 金龙飞等(2022)利用psRNAtarget软件预测出20个靶向拟南芥AtWRI1的候选miRNA (包括miR156、miR164和miR780等), 但这些靶向关系及miRNAs能否在转录后水平靶向WRI1调控油脂合成的研究尚未见报道。本研究通过双荧光素酶报告检测及RNA pull down技术进一步证实了前期预测的沙棘hrh-miRn458靶向转录因子WRI1, 这为非编码RNA直接参与转录因子WRI1调控油脂合成提供了例证, 但hrh-miRn458-WRI1模块通过转录因子WRI1下游靶基因调控油脂合成的分子机制仍有待进一步研究。
本文的其它图/表
|
 首页
首页