|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
植物驱动蛋白: 从微管阵列到生理活动调控
植物学报
2022, 57 (3):
358-374.
DOI: 10.11983/CBB22007
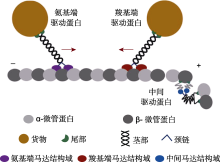
驱动蛋白(kinesin)是以微管为轨道的分子马达, 其催化ATP水解为ADP, 将贮藏在ATP分子中的化学能高效地转化为机械能, 在细胞形态建成、细胞分裂、细胞运动、胞内物质运输和信号转导等多种生命活动中发挥重要作用。对植物驱动蛋白的研究落后于动物和真菌, 其原因不仅由于植物进化出独有的驱动蛋白家族, 而且其家族成员数量远多于动物驱动蛋白。该文主要总结了驱动蛋白在微管阵列动态组织, 包括周质微管和有丝分裂早前期微管带、纺锤体及成膜体中的角色和功能, 以及其对植物生理活动的调控作用。同时对重要经济作物大豆(Glycine max)中的驱动蛋白进行了系统分析、分类及功能预测, 发现大豆驱动蛋白数量庞大。结合公共数据库中大豆转录组数据, 对部分大豆驱动蛋白进行功能预测, 以期对大豆及其它作物驱动蛋白功能研究提供线索和启示。
表1
植物驱动蛋白的主要功能
正文中引用本图/表的段落
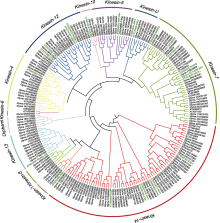
本文对植物驱动蛋白在微管阵列动态组织中的功能及对生理活动的调控作用进行综述(表1)。同时结合公共数据库, 依据基因组织表达特异性和结构的保守性, 对重要经济作物大豆(Glycine max)的驱动蛋白进行了系统分析、分类和功能预测, 以期为大豆等作物驱动蛋白功能研究提供理论依据。
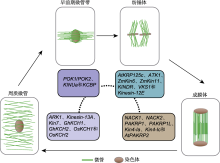
植物细胞分裂间期微管紧贴质膜并与质膜平行排列, 形成二维平面的阵列, 其形成是初始细胞迅速生长的标志和关键步骤(Yuan et al., 1994)。根毛细胞生长是高度动态的过程, 需要微管微丝细胞骨架精准且迅速地转换, 在对根毛细胞骨架的研究中发现了相关驱动蛋白。拟南芥ROP2 (Rho of plant) GTPase控制根毛细胞骨架建成(Fu et al., 2002)。后续研究发现, 拟南芥KINU家族成员MRH2 (morphogenesis of root hair 2)/ARK1 (armadillo-repeat kinesin 1)是控制微管形成的重要组分, 可能参与根毛极性生长期间ROP2 GTPase控制的微管和微丝协同(Jones et al., 2006; Yang et al., 2007; Eng and Wasteneys, 2014)。自身启动子驱动的ARK1-GFP在微管正极大量积累, 回补ark1-1突变体根毛生长缺陷表型, 瞬时过表达 ARK1-GFP增加微管灾变(catastrophe)频率, 表明ARK1通过促进微管灾变调控根毛极性生长(Eng and Wasteneys, 2014)。
拟南芥Kinesin-5家族成员AtKRP125c定位于纺锤体, 并且在其突变体根细胞中可观察到大量纺锤体结构变异(Bannigan et al., 2007), 这些表型与动物及真菌的kinesin-5突变体细胞相似, 表明驱动蛋白Kinesin-5在纺锤体形态建成中功能保守。Kinesin-14家族成员ATK1 (Arabidopsis thaliana kinesin 1)/Kinesin-14A与ATK5/Kinesin-14B均定位于纺锤体, 其突变体均表现纺锤体不同程度的异常。atk1-1突变体在有丝分裂及雄性减数分裂时纺锤体两极均出现无法聚集的现象(Chen et al., 2002; Marcus et al., 2003); 而atk5-1突变体有丝分裂纺锤体出现异常伸长、加宽及频繁蜷曲现象, 而在雄性减数分裂过程中纺锤体基本正常(Ambrose et al., 2005; Ambrose and Cyr, 2007), 表明ATK1在雄性减数分裂中起主要作用, 但ATK1与ATK5调控纺锤体组装的机制仍有待进一步研究。
玉米(Zea mays) Kinesin-14家族多个成员参与纺锤体组装。突变体dv1 (divergent spindle-1)中同源基因ZmKin6与ZmKin11突变导致纺锤体组装异常、花粉活性降低, ZmKin6与ZmKin11在有丝分裂及减数分裂中期影响两极纺锤丝聚集(Higgins et al., 2016)。KINDR (Kinesin driver)是由8个ZmKin11同源基因组成的基因簇, 在减数分裂时其编码的驱动蛋白能够沿纺锤体丝激活新着丝粒位点, 拉着该着丝粒优先传递给卵细胞(Dawe et al., 2018)。VKS1 (Varied-kernel-size phenotype 1)与KINDR碱基序列的相似度为94%, 在胚乳发育早期高度表达, vks1突变体胚乳细胞有丝分裂纺锤体组装异常, 姐妹染色单体无法分离, 最终胞质分裂失败、种子变小(Huang et al., 2019)。
成膜体是植物细胞分裂进行胞质分裂组装细胞板的场所。在细胞分裂后期, 纺锤体逐渐缩短变为伞状结构并向两极聚集(Oh et al., 2012); 同时细胞中部平面逐渐形成2个向外扩张的双环状反向平行重叠微管阵列, 其包围区域称为成膜体。在成膜体扩张过程中, 其内部同时进行细胞板组装, 而驱动蛋白在此过程中发挥重要作用。其中研究得最为深入的是烟草Kinesin-7家族成员NACK1 (NPK1-activating kinesin-like protein 1)及其同源蛋白NACK2。研究表明NPK1 (new protein kinase 1)与成膜体形成有关(Nishihama et al., 2001)。通过酵母双杂交筛选出NPK1激活子NACK1, 亚细胞定位显示NPK1与NACK1共定位于成膜体上, nack1突变体因成膜体无法扩张, 进而子细胞无法形成正常的细胞板, 导致胞质分裂失败(Nishihama et al., 2002)。研究揭示植物中调控成膜体形成的保守通路如下: NACK1和NACK2与NPK1或其它MAPK激酶互作, 通过MAPK级联通路磷酸化下游MAP65 (microtubule-associated proteins 65), 磷酸化状态的MAP65与微管结合能力降低, 促进成膜体微管动态变化(Sasabe and Machida, 2012)。
拟南芥花粉中存在Kinesin-12家族成员PAKRP1 (phragmoplast-associated kinesin 1)及PAKRP1L (phragmoplast-associated kinesin 1 like)与激酶TIO (Two-In-One)互作调控成膜体组装通路。PAKRP1和PAKRP1L单突变体没有表型变化, 可能由于它们的氨基酸序列具有74%的相似度, 从而存在功能冗余; 在双突变体有丝分裂中观察到细胞成膜体无法正常组装, 形成有缺陷的细胞板(Lee and Liu, 2000; Pan et al., 2004; Lee et al., 2007)。研究表明PAKRP1和PAKRP1L也是通过MAPK级联通路实现对成膜体的调控: PAKRP1和PAKRP1L与MAP65-3互作, 将MAP65-3定位到成膜体反向平行微管阵列上(Ho et al., 2011)。前期研究显示, MAP65动物同源蛋白PRC1 (protein required for cytokinesis 1)在有丝分裂后期结合反向平行微管阵列, 并招募Kinesin-4家族成员Xklp1 (xenopus kinesin-4)到周质分裂位点, Xklp1通过抑制微管生长调控纺锤体丝重叠程度进而影响成膜体形成(Bieling et al., 2010; Hu et al., 2011)。由此推测在拟南芥有丝分裂后期, PAKRP1和PAKRP1L先沿微管将MAP65-3运输到周质分裂位点, MAP65-3再招募Xklp1调控成膜体组装。小立碗藓中Kinesin-4家族成员Kin4-Ia与Kin4-Ic参与成膜体微管反向平行重叠以及细胞板形成, 导致Kin4-Ia/Kin4-Ic双突变体微管阵列过度重叠, 从而使成膜体向外扩张速率变慢且微管重叠区变厚, 最终形成有缺陷的细胞板。体外实验表明, Kin4-Ic氨基末端结构域具有解聚微管的功能, 推测Kin4-Ia与Kin4-Ic可能通过解聚微管阵列重叠程度调控成膜体以正确的速率和厚度扩张(de Keijzer et al., 2017)。
近年来, 对FRA1蛋白功能的研究更为细致深入。Kong等(2015)利用具有迅速伸长特征的棉纤维鉴定与细胞伸长相关的驱动蛋白, 研究表明, 棉花中FRA1同源蛋白GhKinesin-4A在伸长的棉纤维中显著富集; 他们还利用拟南芥进一步揭示AtKinesin-4A/ FRA1和AtKinesin-4C在调控细胞壁强度、细胞伸长和各种营养与生殖器官的轴向生长方面存在功能冗余, 同时发现FRA1可能运输非纤维素类物质。FRA1蛋白缺失使囊泡在高尔基体附近积累, 并极大地减少果胶分泌, 导致初生与次生细胞壁均受损, 突变体fra1-5细胞壁厚度仅为野生型的一半(Zhu et al., 2015)。FRA1水稻同源蛋白BC12 (brittle culm 12)缺失使细胞壁成分改变及次生细胞壁中纤维素微纤丝异常沉积, 最终导致植株变脆(Zhang et al., 2010)。IMB4 (importin β4)与FRA1互作, 对其进行时空调控从而使其在运输组装细胞壁相关产物中发挥作用。IMB4通过2种方式调控FRA1: (1) 正在组装细胞壁的初始细胞中, 需要大量活化状态的FRA1运输组装细胞壁相关产物, FRA1与IMB4结合抑制蛋白酶对FRA1的降解, 提高FRA1蛋白的稳定性; (2) 在细胞壁组装完成的成熟细胞中, 不需要过多活性状态的FRA1, 此时IMB4直接阻止FRA1马达结构域与微管结合, 从而使FRA1保持非活性状态(Ganguly et al., 2018, 2020)。最新研究表明, FRA1尾部结构域与CMU (cellulose synthase-microtubule uncoupling)互作, 增强周质微管的横向稳定性, 揭示了周质微管稳定纤维素和基质多糖沉积位点的机制(Ganguly et al., 2020)。
线粒体是细胞制造能量的细胞器。早期研究表明线粒体运动与驱动蛋白类蛋白(kinesin related proteins, KRP)有关, 驱动蛋白类蛋白通过结合线粒体表面微管驱动其运动。拟南芥MKRP1 (mitochondria- targeted KRP1)和MKRP2氨基末端具有线粒体靶向信号。亚细胞定位实验表明, MKRP1和MKRP2与线粒体共定位, 但其驱动线粒体运动的分子机制尚不清楚(Itoh et al., 2001)。通过抗体特异性识别和免疫荧光定位, 发现拟南芥Kinesin-14家族成员ATKP1 (kinesin-like protein 1)特异地定位于线粒体(Ni et al., 2005)。以ATKP1尾部结构域为诱饵, 通过酵母双杂交筛选出线粒体外膜蛋白VDAC3 (voltage-dependent anion channel 3), 二者互作参与种子低温萌发过程的呼吸调节(Yang et al., 2011)。
雄性不育系是重要的育种材料。随着对雄性不育研究的深入, 已挖掘出多个控制雄性育性的驱动蛋白。水稻Kinesin-1家族成员PSS1为组成型表达, 在花药减数分裂期表达量达到峰值, PSS1突变导致花粉活力降低和花药开裂缺陷, 从而引起水稻败育(Zhou et al., 2011)。在拟南芥pss1突变体中观察到染色体联会(synapsis)异常, 雄配子减数分裂时染色体分离异常, 导致花粉数量减少及育性缺陷(Zhou et al., 2011)。减数分裂时同源染色体形成交叉(crossover)导致遗传多样性, 拟南芥中PSS1与WIPs (WPP domain-interacting proteins)/SUNs (Sad1/UNC-84)互作, 控制减数分裂联会与交叉(Duroc et al., 2014)。
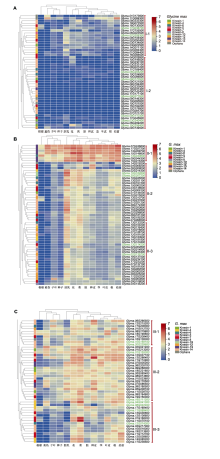
水稻Kinesin-1成员PSS1为组成型表达, 在花药减数分裂时期表达量达到峰值(Zhou et al., 2011)。其大豆同源基因共有4个, 其中Glyma.03G240400、Glyma.08G170500和Glyma.15G256400有表达数据, 均在分支I-2中, Glyma.15G256400在花中表达量最高, 该基因也是分支I-2所有基因中表达量最高的, 推测其在大豆花药减数分裂中具有类似PSS1的功能。拟南芥Kinesin-5家族成员AtKRP125c的同源基因中, Glyma.13G330600 (II-2)相比Glyma.12G074300 (III-3)、Glyma.09G110300 (I-2)和Glyma.15G043400 (无表达数据)在根中优势表达, 表明Glyma.13G330600可能在根细胞有丝分裂纺锤体组装中起重要作用。
值得一提的是, 多个同一驱动蛋白家族基因成对出现, 除了上文提到的Kinesin-10家族仅有的2个成员, Glyma.10G182800和Glyma.20G207200 (II-2), VSK1同源基因Glyma.10G159100和Glyma.- 20G229600 (III-1), Kinesin-4家族FRA1同源基因Glyma.04G041300和Glyma.06G042400, 以及Glyma.14G085200和Glyma.17G239500均在分支III-3中成对出现。Kinesin-7家族Glyma.01G168200 和Glyma.11G075100 (III-3)是在荚中高表达的1对基因, 表明这些基因可能以异源二聚体的形式发挥微管辅助蛋白功能, 其功能与作用机制值得深入研究。
微管是真核生物细胞骨架的重要组分, 在细胞形态建成、细胞分裂、囊泡运输及信号转导等生命活动中发挥重要作用(岳剑茹等, The ROP2 GTPase controls the formation of cortical fine F-actin and the early phase of directional cell expansion during Arabidopsis organogenesis 1 2002 ... 植物细胞分裂间期微管紧贴质膜并与质膜平行排列, 形成二维平面的阵列, 其形成是初始细胞迅速生长的标志和关键步骤(Yuan et al., Importin-β directly regulates the motor activity and turnover of a Kinesin-4 1 2018 ... 近年来, 对FRA1蛋白功能的研究更为细致深入.Kong等( FRA1 kinesin modulates the lateral stability of cortical microtubules through cellulose synthase-microtubule uncoupling proteins 2 2020 ... 近年来, 对FRA1蛋白功能的研究更为细致深入.Kong等(
本文的其它图/表
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 首页
首页