
|
|
||
|
小桐子半胱氨酸蛋白酶家族和相应miRNAs的鉴定及其对低温锻炼的响应
植物学报
2021, 56 (5):
544-558.
DOI: 10.11983/CBB21014
小桐子(Jatropha curcas)是一种极具潜力的能源植物及冷敏植物, 12°C低温锻炼可显著提高其耐冷性。在全基因组水平上对小桐子半胱氨酸蛋白酶家族及靶向其基因的miRNAs进行鉴定、生物信息学分析和表达特性分析, 并对该基因家族成员与miRNAs互作参与调控小桐子对低温锻炼的响应进行解析。结果表明, 在小桐子基因组中共鉴定到39个半胱氨酸蛋白酶基因, 定位于11条染色体上, 可分为6个亚家族(C1A、C2、C12、C13、C14和C15), 编码181-2 158个氨基酸残基的多肽, 均具有Cys和His活性位点。基于miRNA组和降解组测序结果, 发现有283个miRNAs靶向调控小桐子半胱氨酸蛋白酶基因家族的14个成员。对靶向JcDEK1、JcRD21B和JcXBCP3L的miRNAs在12°C低温锻炼过程中的共表达分析表明, 这些miRNAs参与半胱氨酸蛋白酶基因表达的调控, 且这种调控可能与低温锻炼诱导的小桐子耐冷性增强有关。研究结果有助于深入理解小桐子半胱氨酸蛋白酶基因家族功能及其与相应miRNAs的互作, 以及通过互作调控小桐子对低温的响应。  View image in article
图6
小桐子幼苗叶片半胱氨酸蛋白酶家族基因在12°C低温锻炼期间基因表达的热图
正文中引用本图/表的段落
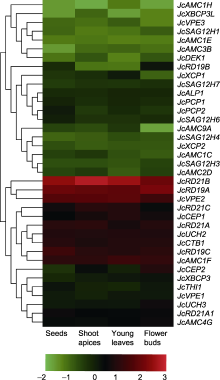
前期研究表明, 12°C低温锻炼48小时可显著增强小桐子在1°C下的耐冷性(Ao et al., 2013a, 2013b)。利用本实验室测序获得的小桐子在低温锻炼过程中的转录组数据(PRJNA661688), 对半胱氨酸蛋白酶家族基因的表达谱进行分析(图6)。结果表明, 该家族大部分成员在12°C低温锻炼期间均明显上调表达, 其中JcAMC1H在6小时表达量最高; 而JcCEP2、JcSAG12H7和JcSAG12H4在低温锻炼前期呈下调表达趋势。
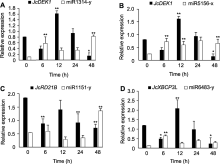
基于前述靶向半胱氨酸蛋白酶家族基因miRNAs的鉴定结果, 利用本实验室在小桐子低温锻炼期间测序获得的miRNA组数据, 挑选40个差异倍数在2.5以上的miRNAs进行数字表达谱分析(图8)。结果表明, miR1314-y、miR7736-y、miR3630-y、miR1039-x和miR5168-x在12°C低温锻炼下表达显著下调, miR1215-x和miR5278-y均在低温锻炼6和12小时后表达显著上调, miR6485-x、novel-m0130-5p、miR8639-y、miR3522-x和miR6112-y在12°C低温锻炼6小时后表达显著上调。值得注意的是, miR1314-y、miR7736-y、miR3630-y、miR1039-y、miR5168-x、novel-m0074-3p和miR5156-x在低温锻炼期间表达明显下调(图8), 而这些miRNAs靶向的半胱氨酸蛋白酶基因JcDEK1在低温锻炼期间表达逐渐上调(图6), 它们之间呈现明显的负相关。与之类似, JcRD21C在低温锻炼期间表达下调(图6), 而靶向其基因的novel-m0113-3p在低温锻炼12-48小时期间表达上调, miR1215-x在低温锻炼6-12小时期间表达上调, miR8639-y、miR3522-y和miR6112-y在低温锻炼6小时后表达明显上调(图8)。这些miRNAs与靶向的JcRD21C总体呈负调控趋势, 表明miRNAs与其靶向的半胱氨酸蛋白酶基因存在负调控关系。
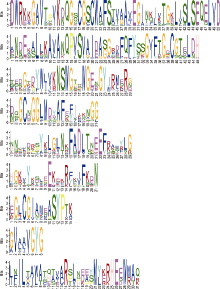
本研究在基因组层面上对小桐子半胱氨酸蛋白酶全基因家族进行了生物信息学分析, 共鉴定到39个家族成员, 可分为C1A、C2、C12、C13、C14和C15共6个亚家族, 定位于11条染色体上, 部分基因之间发生了片段重复, 如JcRD21A和JcRD21A1, JcSAG12H6和JcSAG12H7; 而JcSAG12H5和JcSAG12H4、JcSAG12H3和JcSAG12H4、JcSAG12H5和JcSAG12H3之间的氨基酸同源性达90%以上, 位于不同染色体之间, 推测可能是发生了染色体异位重复(图1, 图4)。蛋白结构域分析结果表明, 小桐子半胱氨酸蛋白酶家族均具有共同的Cys和His活性位点, 保守基序结构域前体序列为ExxxRxxxxxxN/ VxxxNx (图2, 图3)。基于RNA-seq的研究结果表明, 小桐子半胱氨酸蛋白酶基因家族的39个成员在植株的不同组织/器官及在低温锻炼过程中均呈现空间和时间的差异化表达(图5, 图6)。Zou等(2018)研究表明, C1A家族大部分成员在小桐子叶片中表达量较高, 根据本研究结果及与其它5个亚家族成员进行横向比较, 我们发现在种子、花蕾、茎端和叶片中表达量占比较大的基本是C1A亚家族成员, 表明该基因家族不同成员的表达水平在小桐子生长发育的不同阶段及对不同外界环境刺激过程中存在差异化响应。这些结果对我们深入理解小桐子半胱氨酸蛋白酶家族各成员在生长发育过程和对环境变化响应中的作用具有参考价值。
研究证明, 逆境胁迫下细胞内变性或失活蛋白的标记、定向降解及其调控在植物响应和适应逆境胁迫过程中起重要作用, 而半胱氨酸蛋白酶家族作为植物蛋白水解酶的主要组分, 其家族不同成员参与植物多种抗逆调控。例如, 甘蓝型油菜(Brassica napus)在4°C低温下, 其编码半胱氨酸蛋白酶的bcp-15转录本积累最多; 置于4°C的第1天, 转录水平增加了8.5倍, 4天后, 转录水平下降到基础水平的6.4倍(Stroeher et al., 1997)。在PEG、盐和冷胁迫下, 小麦(Triticum aestivum) PLCP基因(TaCP)表达显著上调(Zang et al., 2010)。过表达旱柳(Salix matsudana)半胱氨酸蛋白酶基因SmCP可提高转基因拟南芥的耐盐性(Zheng et al., 2018)。本实验室前期研究表明, 12°C低温锻炼48小时可显著提高小桐子幼苗在1°C低温胁迫下的耐冷性, 这涉及抗氧化防御系统的激活、渗透调节能力增强及膜质脂肪酸组分改变(Ao et al., 2013a, 2013b; Li et al., 2014)。在本研究中, 12°C低温锻炼的早期阶段, JcUCH3、JcRD19C、JcXBCP3、JcUCH2、JcDEK1和JcAMC1H等明显上调表达(图6, 图9), 暗示半胱氨酸蛋白酶基因家族的部分成员可能参与小桐子对低温的早期响应及信号转导过程。但半胱氨酸蛋白酶是一个大家族, 由许多成员组成, 其成员如何差异化地响应植物生长发育的各个阶段及多种环境刺激与胁迫尚不清楚, 值得深入研究。挑选该基因家族中对低温胁迫高响应的基因, 通过基因组编辑、RNA干扰以及过表达等手段进一步进行基因功能鉴定, 将有助于阐明半胱氨酸蛋白酶基因家族特定成员参与植物对低温的响应及调控植物耐冷性的机理。
已有研究表明, 大量miRNAs参与众多生物基因表达的调控(Wani et al., 2020; 郝大海和龚明, 2020; 张翠桔等, 2020), 但miRNAs是否参与调控半胱氨酸蛋白酶家族基因尚未见明确报道。本研究基于小桐子miRNA组和降解组测序结果, 发现有14个半胱氨酸蛋白酶基因受283个miRNAs靶向调控(图7)。比较低温锻炼过程中miRNAs与其靶向的半胱氨酸蛋白酶基因的表达谱(图6, 图8), 发现许多miRNAs与其靶向基因存在负调控关系。对JcDEK1、JcRD21B和JcXBCP3L及靶向它们的miRNAs的qRT-PCR共表达分析结果也表明, 这些半胱氨酸蛋白酶基因与miRNAs之间存在负调控关系, 在低温锻炼12-48小时期间尤为明显(图8), 表明这些miRNAs参与半胱氨酸蛋白酶基因的调控, 且其调控作用与低温锻炼诱导的小桐子耐冷性有关, 但具体调控哪些生化与分子途径还有待深入研究。后续可通过多组学联合分析, 解析基因组中编码半胱氨酸蛋白酶基因的转录因子与miRNA的关系, 有望进一步阐明miRNAs与半胱氨酸蛋白酶基因互作调控基因表达的具体机制, 以及这种互作如何参与调控植物对低温的响应与适应。
本文的其它图/表
|
 首页
首页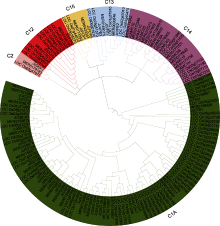



{kind=link}