|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
丁香属次生代谢产物及其与系统演化和地理环境的关联
植物学报
2021, 56 (4):
470-479.
DOI: 10.11983/CBB20178
环境塑造的植物次生代谢产物富于变化, 也可能带有系统演化的信息。由于完整或具有系统学代表性的专属植物收集存在较大困难, 使得次生代谢产物与系统学的关联研究尚不多见。通过文献汇总获得了存在于丁香属(Syringa)植物根、茎、叶和花中的10类377个次生代谢产物, 主要涉及甲戊二羟酸途径、脱氧木酮糖磷酸酯途径以及莽草酸途径。在叠加丁香属的系统演化背景后发现: 在先分化的组系中特定类型次生代谢产物的优势度较高, 后继分化的组系成分优势度降低, 化学多样性呈增加趋势, 各类次生代谢产物的相对占比趋于均衡; 苯丙素类和环/裂环烯醚萜类化合物的表达具有明显的系统保守性。在叠加了地理分布跨度后发现: 部分后继分化的局域种比在先分化的广布种具有更为多样的次生代谢成分; 木脂素类成分的占比优势与环境胁迫相关。该文为化学多样性与进化的关联研究及次生代谢调控的系统性研究提供了新的启示。
表1
已报道次生代谢成分的丁香属种质地理分布
正文中引用本图/表的段落
丁香属植物含有丰富的次生代谢产物, 具有相同母核的同类成分常因取代基团类型、数量、位置和空间构象的不同而数量众多。经整理我们发现已报道的次生代谢产物共有377个(附表1), 按其母核结构可分为10类: 木脂素类成分74个, 环/裂环烯醚萜类成分118个, 倍半萜类成分67个, 三萜类成分34个, 单萜类成分3个, 黄酮类成分13个, 苯丙素类成分59个, 脂肪酸类3个, 醌类2个, 生物碱类4个。其中含量较高的成分多是特定功能的重要贡献者, 其对应的代谢途径也是我们分析的重点。由于针对属内组系水平的化学多样性分析的客观性取决于特定物种的研究案例数量和属下分类等级上的物种覆盖度, 我们选取的文献所述物种在丁香属内涉及5个组系(表1)。在这些文献中, 中国原产的单种系羽叶丁香系(Ser. Pinnatifoliae) 21篇; 东亚和欧洲分布的欧丁香系(Ser. Syringa) 20篇, 其中东亚广布种紫丁香(S. oblata)和朝阳丁香(S. oblata var. dilatata)分别为10篇和3篇, 东南欧洲广布种欧洲丁香(S. vulgaris) 6篇, 中国西北-中西亚分布种阿富汗丁香(S. afghanica) 1篇; 东亚及远东广布的短花冠管组(Sect. Ligustrina)中的暴马丁香(S. amurensis) 6篇和日本丁香(S. reticulata) 3篇; 中国华北和东北分布的巧玲花系(Ser. Pubescentes)共6篇, 其中包括华北分布种巧玲花(S. pubescens)和东北分布种关东丁香(S. velutina)各3篇; 中国西南和东北亚分布的红丁香系(Ser. Villosae) 2篇, 其中包括西蜀丁香(S. komarowii)和辽东丁香(S. wolfii)各1篇。我们分别在组系(表2)和物种(表3) 2个分类水平上对次生代谢产物进行归类整理。通过python+seaborn软件包(
- 表示该类产物在特定物种中未见报道。成分来自根、茎、花、叶和茎皮。各类成分计数包括具有相同母核但取代基团、基团数量、基团位置和空间构象不同的化合物。阿富汗丁香、辽东丁香和西蜀丁香数据分别仅来源于1篇文献。
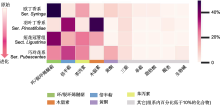
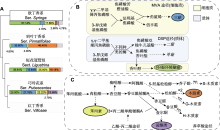
(A) 丁香属组系的系统演化关系(Li et al., 2012)及各类产物在各组系中的占比; (B) 甲戊二羟酸途径(MVA)和脱氧木酮糖磷酸酯途径(DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) 莽草酸途径(刘津等, 2016; 张旭等, 2019)。图(A)组系下方的色条表示各类产物(颜色含义同
本研究表明, 丁香属在组系分化的先后梯度上表现为成分的优势度降低而化学多样性水平升高。在先分化的组系有更具优势的特定类型代谢产物, 而后续分化的组系特定类型成分的优势度降低而多样性水平升高(图1)。首先分化的欧丁香系物种虽有苯丙素、木脂素、黄酮、单萜、倍半萜、三萜和脂肪酸类等众多类型的代谢产物, 但丝毫不影响环/裂环烯醚萜类产物的绝对占比优势(高达57.14%)。这使得紫丁香和欧洲丁香能在更多样的环境选择压力下生存, 包括由经纬度和海拔的较大变幅所造成的气温、光照以及土壤环境的巨大差异, 欧丁香系中的欧洲丁香能够在巴尔干半岛南部山区中广泛分布, 紫丁香也能在中国北方的广阔区域中繁衍生息(表1, 表3)。对于较先分化的羽叶丁香系, 相对占比高达46.40%的木脂素同样具有优势。在此后持续的演化分异中, 特定产物的优势出现降低趋势。如果说处在进化中间位置的短花冠管组尚有37.64%的环/裂环烯醚萜类成分占比, 相对于较先分化的羽叶丁香系已经明显降低的话, 那么后续分化的巧玲花系很难看出特定化合物的绝对优势, 其中环/裂环烯醚萜类(25.00%)、倍半萜类(27.78%)和苯丙素类(22.22%)成分的数量占比则更为均衡(图1)。
丁香属中的环/裂环烯醚萜类和苯丙素类产物呈现较强的系统遗传保守性, 甚至遥远的地理隔离也未造成近缘种优势代谢产物的明显差异。紫丁香和欧洲丁香是系统学意义上的姊妹系(Li et al., 2012), 虽然二者在自然分布上存在着东亚与巴尔干半岛的遥远亚欧间断分布(表1), 却均具有丰富的环/裂环烯醚萜类和苯丙素类产物(表3)。苯丙素类产物形成的保守性还体现在短花冠管组的2个近缘种暴马丁香和日本丁香中。暴马丁香广泛分布于中国北方及远东地区, 日本丁香仅在日本呈局域分布, 但2个系统学的近缘种同样具有占比相近的苯丙素类成分(表1, 表3)。由此表明, 环/裂环烯醚萜类和苯丙素类成分的形成具有较强的遗传保守性, 强烈的保守性没有因遥远的地理距离而消弱。与此同时, 在相邻的近缘组系中出现了较高占比的特定类别化合物的连续分布(图1)。图2A显示, 木脂素以较高的占比出现在相邻的羽叶丁香系(46.40%)和短花冠管组(18.18%)中; 倍半萜分别以36.80%、13.64%和27.78%的较高占比出现在相邻的羽叶丁香系、短花冠管组和巧玲花系中; 苯丙素以25.00%和22.22%的占比出现在相邻的短花冠管组和巧玲花系中; 环/裂环烯醚萜类分别以38.64%和25.00%的较高占比出现在相邻的短花冠管组和巧玲花系中。值得一提的是, 在针对同一个物种的诸多文献进行化合物汇总时, 我们发现欧洲丁香的样品取自罗马尼亚(Varga et al., 2019)和波兰(Filipek et al., 2019)多地, 但这些来自不同环境的同一物种却具有高度相似的次生代谢产物。这表明在丁香属特定的次生代谢途径中确实存在无法被环境异质性改变的系统学保守性。
在分别从系统演化和地理环境角度讨论丁香属次生代谢产物的分布趋势之后, 需要面对的问题还有代谢背景下的系统演化与地理分布跨度的关系。研究表明, 紫丁香和欧洲丁香是丁香属系统学原始组系中的2个近缘姊妹系物种(Li et al., 2012), 它们也分别是东亚和欧洲的广布种(张美珍等, 1992; Fiala, 2008), 而后续分化的红丁香系(Li et al., 2012)中的大多数物种都属于生态幅狭窄的局域种。已知温度和水分是限制物种扩散的主要环境因子(高坤等, 2018; 魏华等, 2018), 丁香地理分布范围的差异一定程度上由对温度和水分敏感的光合同化产物形成和水分利用效率等初生代谢机制的差别决定(Cui et al., 2016)。紫丁香和欧洲丁香的广布表明其初生代谢机制保证了它们能够适应东亚和东南欧洲的气候变化, 同时强大的系统学保守性使相距万里的2个系统学意义上的近缘姊妹种均具有环/裂环烯醚萜和苯丙素类产物(表1, 表3), 使它们有能力通过这两类优势次生代谢产物的形成, 增强对小环境中生物及非生物扰动的适应能力, 从而改善植株的生长状况并实现高效的繁衍生息(Konno et al., 1999; 刘盟盟等, 2017; Sharma et al., 2019)。而对于性喜冷凉的红丁香系中的红丁香和匈牙利丁香2个近缘种, 气孔较大且密度较低的叶片表皮特征很可能使其在初生代谢热量耗散和叶片水分保持能力上存在限制(高艳等, 2008), 因此其分布局限在高海拔的冷凉或邻水生境(张美珍等, 1992; Lendvay et al., 2016)而成为地理分布上的局域种。在这类水热条件适宜的小环境中, 这些较晚分化的局域种不必应对地域扩散中温度和水分的巨大变化, 有条件形成更加多样的次生代谢产物, 在适宜的生境中最大限度地改善生长质量。这种“初生代谢决定生存格局, 次生代谢改善生活质量”(刘晓侠等, 2015)的情况也发生在同样进化却局域分布的巧玲花系中(图1)。
附表1 丁香属已发表的次生代谢产物信息汇总
本文的其它图/表
|
 首页
首页