|
|
||||||||||||||||||||||||||||||
|
类受体激酶FER调节植物与病原菌相互作用的分子机制
植物学报
2021, 56 (3):
339-346.
DOI: 10.11983/CBB20180
植物细胞依赖细胞质膜上的受体感知并传递环境信号, 而受体通过与配体特异结合启动一系列下游信号转导途径, 维持植物正常的生命活动及其对外界环境变化的适应。类受体激酶是其中一类重要受体, 通常由胞外结合结构域、跨膜结构域和胞内激酶结构域3部分组成, 是植物适应外界环境变化的重要调节枢纽。FER属于CrRLK1L类受体蛋白激酶家族, 最早被发现在高等植物雌雄配子体识别过程中发挥作用。随后, 众多研究表明, FER在植物生长发育、激素间交互作用、植物与病原菌互作和逆境响应等多种生物学过程中扮演重要角色, 是近年来植物信号通路研究领域的“明星蛋白”。随着植物病理学研究的不断深入, FER在植物与病原菌互作过程中的功能备受关注。该文主要综述FER调节植物与病原菌互作的研究进展, 旨在为进一步解析类受体蛋白激酶在植物细胞响应病原菌侵染过程中的信号转导机制提供参考。
表1
FER调节植物细胞与病原菌间的相互作用
正文中引用本图/表的段落
FER属于长春花类受体激酶家族(Catharanthus roseus RLK1-like kinases, CrRLK1Ls) (Escobar-Restrepo et al., 2007), 最早被发现能够介导雄配子体和雌配子体之间的识别(Huck et al., 2003; Escobar- Restrepo et al., 2007)。拟南芥中的FER与该家族的其它16个蛋白在类受体激酶超家族中形成1个分支, 其胞外结构域中的Malectin-like蛋白基序具有结合二聚及寡聚葡萄糖的能力(Li and Zhang, 2014), 暗示该类受体激酶在植物细胞间互作及植物-环境互作过程中发挥重要作用。近年研究表明, FER广泛参与调节高等植物的受精、细胞伸长、生物及非生物胁迫响应等不同的生物学过程(Chen et al., 2020; Ji et al., 2020b; Zhang et al., 2020b)。FER是胚囊接受花粉管所必需, 并且是雌配子体育性的重要调节因子(Huck et al., 2003)。拟南芥fer突变体中雌配子体对花粉管接收发生功能缺陷, 雌雄配子之间的相互作用被破坏, 导致花粉管生长失控, 不能破裂释放精细胞, 最终导致育性下降(Huck et al., 2003)。进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(Escobar-Restrepo et al., 2007)。Duan等(2020)研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊。花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节。NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE。有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞。阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(Yu et al., 2020)。
另外, FER在果实成熟过程中也发挥重要作用。在草莓(Fragaria × ananassa)果实中过表达FER- like蛋白FaMRLK47能延迟草莓果实的成熟, 下调FaMRLK47表达则加速果实成熟(Jia et al., 2017a)。异源表达苹果(Malus × domestica) FER-like蛋白MdFERL1/MdFERL6可抑制番茄(Solanum lycoper-sicum)果实产生乙烯, 并延缓果实成熟, 而MdFERL6在苹果愈伤组织中过表达抑制乙烯产生(Jia et al., 2017b)。番茄SlFERL在果实中表达量较高, 受果实成熟相关转录因子RIN和TAGL1共调控; Ji等(2020a)通过表面等离子共振结合质谱分析技术鉴定到若干可能与SlFERL互作的蛋白, SlFERL能与乙烯合成途径的关键酶SlSAMS1互作, 调节乙烯的生物合成, 进而影响果实的成熟进程(Ji et al., 2020a)。
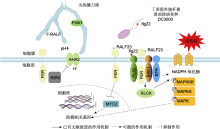
近年来, FER调节植物与病原菌间互作的报道不断涌现(表1)。Keinath等(2010)以FLS2的配体flg22处理15N/14N标记的拟南芥悬浮细胞, 采用定量蛋白质组学技术, 量化flg22刺激后15N/14N标记的拟南芥细胞质膜的早期反应, 重点分析了DRM (detergent- resistant membranes)组分中蛋白表达水平的变化。 结果表明, 经flg22刺激后, 细胞质膜上DRM组分富集的蛋白种类发生显著变化, 15分钟内64个蛋白在DRM组分中显著富集, 其中转运体和RLKs的富集程度最为显著(包括FLS2 (FLAGELLIN-SENSING 2)和FER)。随后该研究组采用反向遗传学和药理干预方法分析了这些蛋白在flg22诱导的防御反应中的作用, 结果表明, 3个候选突变体(det3、aha1和fer4)在flg22 诱发的活性氧积累、丝裂原活化蛋白激酶(mitogen- activated protein kinase, MAPK)活性和气孔关闭等方面表现出功能缺陷, 暗示这3种蛋白可能是调节植物先天免疫的新组分(Keinath et al., 2010)。
此外, FER也负调控寄主植物对尖孢镰刀菌(Fu- sarium oxysporium)的免疫应答。Masachis等(2016)将尖孢镰刀菌接种到番茄根部, 分别供给水、pH5.0和pH7.0的缓冲液, 发现在pH5.0条件下, 植物的死亡率相比水和pH7.0缓冲液条件下更低, 说明胞外碱化促进尖孢镰刀菌对寄主植物的侵染。有研究表明, 尖孢镰刀菌中的丝裂原活化蛋白激酶Fmk1对尖孢镰刀菌的侵染至关重要, 胞外pH值从5.0上升至7.0可触发Fmk1快速磷酸化, 激活与致病性相关的MAPK信号级联反应, 从而促进侵入性菌丝生长和病原菌致病(Masachis et al., 2016)。进一步研究发现, 尖孢镰刀菌会产生一种类似于植物分泌的小肽F-RALF, 这种小肽与RALF1类似, 可诱导宿主组织碱化, 从而增强尖孢镰刀菌对寄主植物的侵染效率(Masachis et al., 2016)。拟南芥突变体fer4对尖孢镰刀菌产生的F-RALF不敏感, 与野生型相比其对尖孢镰刀菌菌株表现出更强的抗性, 暗示FER负调控寄主植物对尖孢镰刀菌的免疫应答。该研究揭示了一种植物病原真菌干扰植物正常生理功能的新机制, 即F-RALF与FER结合, 阻断AHA2 (H+-ATPase 2)介导的H+外流, 提高根系细胞外pH值, 促进病原菌侵染, 从而负调控免疫反应(Masachis et al., 2016)。
另外, FER也起到类似支架蛋白的作用, 介导植物免疫应答反应。Stegmann等(2017)研究发现, 拟南芥SITE-1蛋白酶(S1P)剪切内源性RALF前体肽, FER抑制植物免疫反应。研究表明, 在elf18、flg22和几丁质的作用下, s1p-3和s1p-6单突变体产生更多的ROS, 对低致病力的丁香假单胞菌番茄致病变种(Pseudomonas syringae pv. tomato) (Pto) DC3000 COR- (coronatine-minus)表现出更高抗性, 因此证明S1P是免疫负调控因子(Stegmann et al., 2017)。过表达RALF23能够抑制elf18触发的ROS生成, 增强植株对Pto DC3000 COR-和黄瓜枯萎菌Plectosp- haerella cucumerina的敏感性, 而RALF23缺失促进elf18触发的ROS产生, 增强植物对Pto DC3000的抗性, 表明RALF23同样负调控免疫。进一步研究显示, fer2和fer4突变体对elf18、flg22和几丁质的敏感性较低, 对Pto DC3000 COR-更敏感, 说明FER能正向调节免疫过程。flg22处理后, FER与FLS2和BAK1之间的互作增强, 而在fer4突变体中, flg22诱导的FLS2- BAK1复合物组装减少, fer4/FER-GFP转基因株系中复合物的水平恢复正常。RALF23过表达对flg22诱导的FLS2-BAK1复合物形成也有类似影响。值得注意的是, RALF23或FER的缺失并不影响FLS2、EFR或BAK1的积累。上述结果表明, FER促进配体诱导的FLS2/EFR与其共受体BAK1之间形成复合物, 而RALF23抑制这一过程, 进一步证实FER是调节免疫受体复合物形成的重要支架组分, 与其受体和共受体共同定位于质膜微区中(Stegmann et al., 2017)。这种FER-RALF模块可能通过调节不同的受体激酶复合物, 在生长发育或环境感知等过程中发挥重要作用(Stegmann et al., 2017)。Huang等(2020a)研究表明, 水稻中类FER受体FLR (FER-like receptor)家族成员能够调节水稻与稻瘟病菌(Magnaporthe oryzae)互作。他们利用CRISPR/Cas9基因编辑技术成功敲除了FLR家族的14个成员, 结果显示FLR1和FLR13的突变体对稻瘟病的敏感性增强, 而FLR2和FLR11的功能缺失增强了水稻对稻瘟病的抗性, 且几乎未对水稻生长造成影响(Huang et al., 2020a)。
F-RALF, MYC2, FLS2, EFR, BAK1, RALF23 and MAPK see To grow or not to grow: FERONIA has her say 1 2014 ... FER属于长春花类受体激酶家族(Catharanthus roseus RLK1-like kinases, CrRLK1Ls) ( The malectin-like receptor-like kinase LETUM1 modulates NLR protein SUMM2 activation via MEKK2 scaffolding 1 2020 ... 值得注意的是, CrRLK1L家族的其它成员在植物与病原菌互作过程中也具有类似功能.LET1 (LETUM- 1)和LET2均是定位于细胞质膜的CrRLK1L家族成员, 其中LET1与MEKK2和NLR蛋白SUMM2形成异源三聚体, MEKK2负责维持LET1和SUMM2的稳定性.SUMM2降解依赖CPR1介导的泛素-蛋白酶体降解途径, 而MEKK2能够抑制这一过程并稳定SUMM2蛋白, 从而诱导免疫自激活和细胞死亡; LET2具有类似的功能, 且能够磷酸化LET1.LET1/2蛋白的质膜定位依赖LLG1, 三者共同参与调控SUMM2介导的免疫自激活和细胞死亡途径, 暗示LLG1-CrRLK1L模块在植物免疫过程中的功能可能广泛存在且很保守( FERONIA receptor kinase interacts with S-adenosylme- thionine synthetase and suppresses S-adenosylme-thio- nine production and ethylene biosynthesis in Arabidopsis 1 2015 ... 此外, FER还可调节植物许多其它生物学过程.FER通过调节RHO GTPase信号调控根毛发育.例如, A fungal pathogen secretes plant alkalinizing peptides to increase infection 5 2016 ... 此外, FER也负调控寄主植物对尖孢镰刀菌(Fu- sarium oxysporium)的免疫应答.
本文的其它图/表
|
 首页
首页