
|
|
||
|
植物果胶的生物合成与功能
植物学报
2021, 56 (2):
191-200.
DOI: 10.11983/CBB20179
果胶作为植物细胞壁多糖之一, 其结构和功能非常复杂。果胶主要由同型半乳糖醛酸聚糖(HG)、鼠李半乳糖醛酸聚糖I (RGI)和鼠李半乳糖醛酸聚糖II (RGII)组成。果胶类成分在维持细胞壁结构的完整性以及细胞间黏附和信号转导等方面发挥重要作用。研究果胶类成分的结构、分布和功能, 对理解细胞壁高级结构的构建和功能具有重要意义。然而, 3种果胶组分在细胞壁内如何交联形成高级结构并发挥生物学功能, 目前尚不明确。该文重点阐述果胶3种组分(HG、RGI和RGII)的生物合成、功能以及果胶的显微成像, 旨在为植物果胶结构及功能研究提供参考。 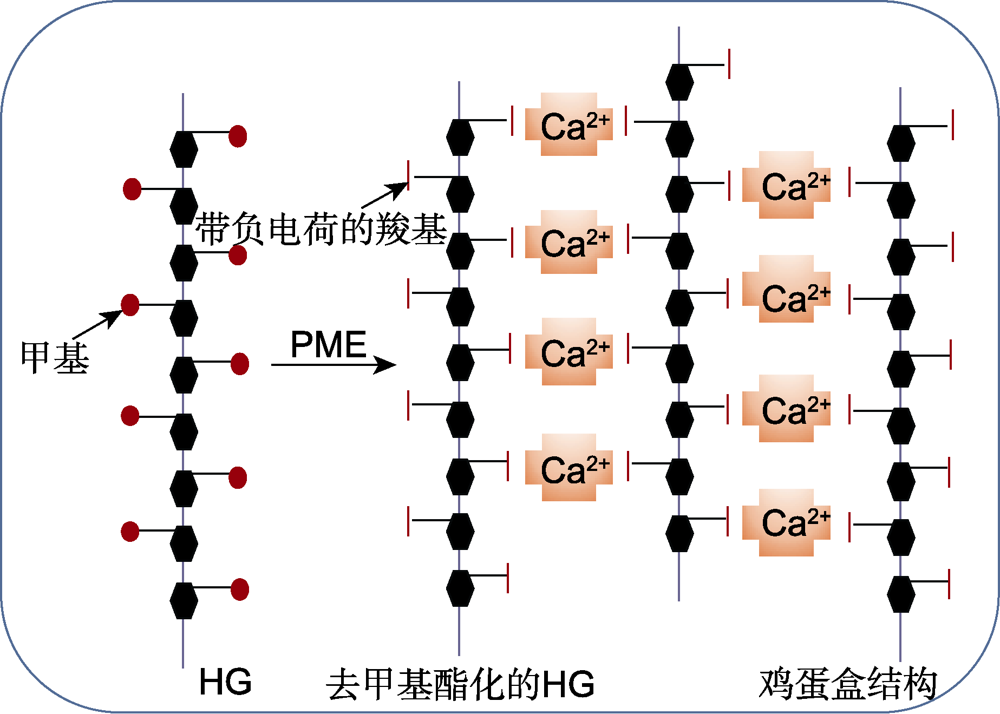 View image in article
图1
同型半乳糖醛酸聚糖(HG)多糖与Ca2+交联的“鸡蛋盒”模型(改自Micheli,
高甲基酯化的HG被果胶甲基酯酶(PME)去甲基酯化, 产生带负电荷的羧基。在Ca2+存在时, 2条HG链的羧基与Ca2+相互作用, 形成“鸡蛋盒”结构。
正文中引用本图/表的段落
HG在高尔基体中以高甲基酯化的形式合成, 随后由果胶甲基酯酶(pectin methylesterase, PME)选择性地去甲基酯化(Caffall and Mohnen, 2009), 同时释放甲醇和质子(查笑君等, 2010), 生成带负电荷的羧基(Wormit and Usadel, 2018)。去甲基酯化的HG可以增强细胞壁的刚性。例如, PMEs催化HG去甲基酯化后可形成连续的GalAs, 在Ca2+存在时, 带负电荷的羧基与Ca2+相互作用, 形成“鸡蛋盒” (Egg Box Model)结构(图1)。该结构可以通过增强细胞壁刚性来维持细胞壁的完整性(Micheli, 2001; Braccini and Peréz,2001); 花粉管的顶端区域几乎只存在甲基酯化的果胶, 外源施加PME时可诱导其尖端细胞壁增厚, 从而抑制花粉管生长(Bosch et al.2005)。此外, PME去甲基酯化HG的过程中会导致局部pH降低, 激活多种细胞壁降解酶, 从而促进细胞壁的扩张, 使其弹性增强。相反, 当植物体缺少PME时, 细胞壁的弹性降低。研究发现, 果胶甲基酯酶6 (PME6)在保卫细胞中表达量较高, 野生型保卫细胞富含未酯化的HG, 而突变体pme6-1的保卫细胞壁富含甲基酯化的果胶, 其气孔开启/关闭动态范围缩小, 表明异常的果胶甲基酯化改变了保卫细胞的机械特性, 导致气孔无法根据环境信号进行适度的开放与关闭(Amsbury et al.2016)。
Hypermethylated HG is de-methylated by pectin methylesterase (PME) to produce negatively charged carboxyl groups. In the presence of Ca2+, the carboxyl groups of the two HG chains interact with Ca2+ to form an “egg box” structure.... A domain swap approach reveals a role of the plant wall-associated kinase 1 (WAK1) as a receptor of oligogalacturonides 1 2010 ... 细胞壁作为抵御病原体的屏障, 其完整性为植物防御病原体所必需(Hamann, The structure, function, and biosynthesis of plant cell wall pectic polysaccharides 1 2009 ... HG在高尔基体中以高甲基酯化的形式合成, 随后由果胶甲基酯酶(pectin methylesterase, PME)选择性地去甲基酯化(Caffall and Mohnen, Pectic arabinan side chains are essential for pollen cell wall integrity during pollen development 1 2014 ... RGI侧链包括β-1,4-半乳聚糖、阿拉伯聚糖或阿拉伯半乳聚糖, 其组成和结构影响细胞壁的强度、弹性和柔韧性等机械性能.例如, 侧链阿拉伯聚糖和阿拉伯半乳聚糖的长度和数量部分决定初生细胞壁的相对灵活性, 而半乳聚糖含量增加有助于增强细胞壁的硬度和刚度(Ulvskovet al. Synthetic methyl hexagalacturonate hapten inhibitors of anti-homogalacturonan monoclonal antibodies LM7, JIM5 and JIM7 2 2003 ... 在果胶多糖的3种组分中, 对于HG的标记成像研究较多, 用于标记HG的单克隆抗体有JIM5、JIM7和2F4等.其中, JIM5的最佳识别表位可能是4个以上相邻的未酯化GalA残基, 这些残基与甲基酯化的GalA残基相邻或在其两侧(Clausen et al.
本文的其它图/表
|
 首页
首页
{kind=link}