
收稿日期: 2019-10-30
录用日期: 2019-12-11
网络出版日期: 2019-12-13
基金资助
国际自然科学基金(31670186)
Detection of SUMOylation in Plants
Received date: 2019-10-30
Accepted date: 2019-12-11
Online published: 2019-12-13
曲高平 , 金京波 . 植物蛋白SUMO化修饰检测方法[J]. 植物学报, 2020 , 55(1) : 83 -89 . DOI: 10.11983/CBB19213
SUMOylation, a post-translational modification, is essential for plant normal growth and development. To date, thousands of possible SUMO substrates have been identified, but due to the relatively low SUMOylation level, biological significance of the SUMOylation remains largely unknown. Here, we summarized the SUMOylation detection methods, including in vitro and in vivo SUMOylation assays, which help to understand the functions of SUMOylation in plants.
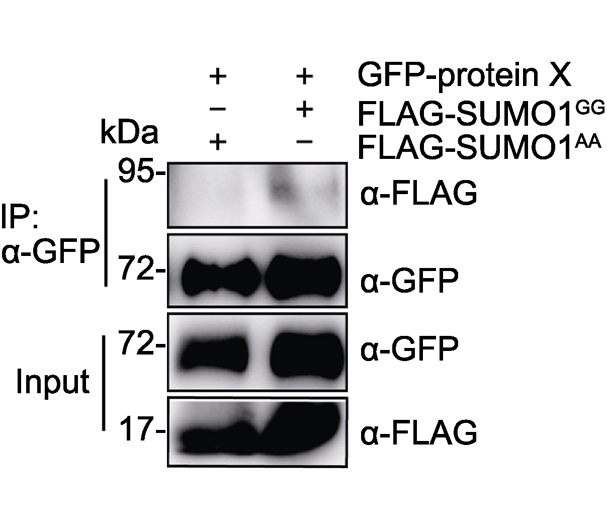
Key words: Arabidopsis; SUMO; SUMOylation; detection methods
| [1] | 徐庞连, 曾棉炜, 黄丽霞, 阳成伟 (2008). 植物SUMO化修饰及其生物学功能. 植物学通报 25, 608-615. |
| [2] | Bernier-Villamor V, Sampson DA, Matunis MJ, Lima CD (2002). Structural basis for E2-mediated SUMO conjugation revealed by a complex between ubiquitin-conjugating enzyme Ubc9 and RanGAP1. Cell 108, 345-356. |
| [3] | Catala R, Ouyang J, Abreu IA, Hu YX, Seo H, Zhang XR, Chua NH (2007). The Arabidopsis E3 SUMO ligase SIZ1 regulates plant growth and drought responses. Plant Cell 19, 2952-2966. |
| [4] | Colby T, Matth?i A, Boeckelmann A, Stuible HP (2006). SUMO-conjugating and SUMO-deconjugating enzymes from Arabidopsis. Plant Physiol 142, 318-332. |
| [5] | Conti L, Price G, O'Donnell E, Schwessinger B, Dominy P, Sadanandom A (2008). Small ubiquitin-like modifier proteases OVERLY TOLERANT TO SALT1 and -2 regulate salt stress responses in Arabidopsis. Plant Cell 20, 2894-2908. |
| [6] | Elrouby N, Coupland G (2010). Proteome-wide screens for small ubiquitin-like modifier (SUMO) substrates identify Arabidopsis proteins implicated in diverse biological processes. Proc Natl Acad Sci USA 107, 17415-17420. |
| [7] | Hermkes R, Fu YF, Nürrenberg K, Budhiraja R, Schmelzer E, Elrouby N, Dohmen RJ, Bachmair A, Coupland G (2011). Distinct roles for Arabidopsis SUMO protease ESD4 and its closest homolog ELS1. Planta 233, 63-73. |
| [8] | Ishida T, Yoshimura M, Miura K, Sugimoto K (2012). MMS21/HPY2 and SIZ1, two Arabidopsis SUMO E3 ligases, have distinct functions in development. PLoS One 7, e46897. |
| [9] | Kong XX, Luo X, Qu GP, Liu P, Jin JB (2017). Arabidopsis SUMO protease ASP1 positively regulates flowering time partially through regulating FLC stability. J Integr Plant Biol 59, 15-29. |
| [10] | Lin XL, Niu D, Hu ZL, Kim DH, Jin YH, Cai B, Liu P, Miura K, Yun DJ, Kim WY, Lin RC, Jin JB (2016). An Arabidopsis SUMO E3 ligase, SIZ1, negatively regulates photomorphogenesis by promoting COP1 activity. PLoS Genet 12, e1006016. |
| [11] | Liu LP, Jiang Y, Zhang XM, Wang X, Wang YB, Han YZ, Coupland G, Jin JB, Searle I, Fu YF, Chen FL (2017). Two SUMO proteases SUMO PROTEASE RELATED TO FERTILITY1 and 2 are required for fertility in Arabidopsis. Plant Physiol 175, 1703-1719. |
| [12] | Miller MJ, Barrett-Wilt GA, Hua ZH, Vierstra RD (2010). Proteomic analyses identify a diverse array of nuclear processes affected by small ubiquitin-like modifier conjugation in Arabidopsis. Proc Natl Acad Sci USA 107, 16512-16517. |
| [13] | Murtas G, Reeves PH, Fu YF, Bancroft I, Dean C, Coupland G (2003). A nuclear protease required for flowering-time regulation in Arabidopsis reduces the abundance of SMALL UBIQUITIN-RELATED MODIFIER conjugates. Plant Cell 15, 2308-2319. |
| [14] | Rodriguez MS (2016). SUMO: Methods and Protocols . New York: Humana Press. |
| [15] | Rytz TC, Miller MJ, McLoughlin F, Augustine RC, Marshall RS, Juan YT, Charng YY, Scalf M, Smith LM, Vierstra RD (2018). SUMOylome profiling reveals a diverse array of nuclear targets modified by the SUMO ligase SIZ1 during heat stress. Plant Cell 30, 1077-1099. |
| [16] | Sadanandom A, ádám é, Orosa B, Viczián A, Klose C, Zhang CJ, Josse EM, Kozma-Bognár L, Nagy F (2015). SUMOylation of phytochrome-B negatively regulates light- induced signaling in Arabidopsis thaliana. Proc Natl Acad Sci USA 112, 11108-11113. |
| [17] | Saleh A, Withers J, Mohan R, Marqués J, Gu YN, Yan SP, Zavaliev R, Nomoto M, Tada Y, Dong XN (2015). Posttranslational modifications of the master transcriptional regulator NPR1 enable dynamic but tight control of plant immune responses. Cell Host Microbe 18, 169-182. |
| [18] | Saracco SA, Miller MJ, Kurepa J, Vierstra RD (2007). Genetic analysis of SUMOylation in Arabidopsis: conjugation of SUMO1 and SUMO2 to nuclear proteins is essential. Plant Physiol 145, 119-134. |
| [19] | Tomanov K, Zeschmann A, Hermkes R, Eifler K, Ziba I, Grieco M, Novatchkova M, Hofmann K, Hesse H, Bachmair A (2014). Arabidopsis PIAL1 and 2 promote SUMO chain formation as E4-type SUMO ligases and are involved in stress responses and sulfur metabolism. Plant Cell 26, 4547-4560. |
| [20] | Yates G, Srivastava AK, Sadanandom A (2016). SUMO proteases: uncovering the roles of deSUMOylation in plants. J Exp Bot 67, 2541-2548. |
| [21] | Yoo SD, Cho YH, Sheen J (2007). Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat Protoc 2, 1565-1572. |
| [22] | Zhao Q, Xie YB, Zheng YY, Jiang S, Liu WZ, Mu WP, Liu ZX, Zhao Y, Xue Y, Ren J (2014). GPS-SUMO: a tool for the prediction of SUMOylation sites and SUMO-interaction motifs. Nucleic Acids Res 42, W325-W330. |
/
| 〈 |
|
〉 |

 首页
首页