|
|
|||||||||
|
拟南芥AtFTCD-L参与根系响应土壤紧实度的机制
植物学报
2025, 60 (4):
551-561.
DOI: 10.11983/CBB24154
植物根系在生长发育过程中响应各种非生物胁迫, 包括干旱、重金属、盐、冷、热以及生理性缺素等, 其中土壤结构特别是土壤紧实度会影响根系的生长与形态, 进而影响作物产量。高尔基体通过囊泡分泌参与根系的生长以及响应非生物胁迫。然而, 高尔基体如何参与根系响应土壤紧实度的机制还不清楚。前期研究发现拟南芥(Arabidopsis thaliana) AtFTCD-L定位在高尔基体反面(trans Golgi network, TGN)上, 参与囊泡的分选和/或分泌, 调节根冠外周细胞中的黏液成分。在前期研究的基础上, 模拟土壤高紧实度生长条件, 观察稳定表达PINs-GFP的纯合体拟南芥植株表型, 通过观察生长素相关荧光信号, 发现AtFTCD-L突变体根尖以及根尖细胞在纵向上短于野生型等材料, 而在横向上宽于野生型等材料, 并且细胞形态明显异常。通过对PINs相关材料进行荧光信号收集, 发现突变体植株中PIN7低表达或不表达。综上表明, AtFTCD-L在拟南芥植株根系中通过调节PIN7的分布或表达来响应土壤紧实度。研究结果为揭示植物根系响应土壤紧实度胁迫的适应机制提供了理论指导。
表1
qRT-PCR引物序列
正文中引用本图/表的段落
选择生长在1/2MS培养基上与梯度琼脂培养基中的野生型拟南芥根, 称取相同重量后, 利用Trizol试剂提取RNA。参考反转录试剂盒使用说明书, 将RNA反转录为cDNA。通过基因上下游引物进行实时荧光定量PCR, 检测AtFTCD-L基因在不同阻力条件下的表达情况, 以Actin8为内参基因。引物序列见表1。采用Excel软件处理实验数据, 采用2-ΔΔCT法分析基因的相对表达量(Livak and Schmittgen, 2001)。
将稳定表达PIN7-GFP的野生型、突变体与回复突变体等植物材料分别点播到梯度琼脂培养基和普通1/2MS培养基上。对梯度培养基中生长7天的拟南芥根尖进行表型观察与统计, 对1/2MS培养基上生长5天的拟南芥根尖进行Confocal观察, 统计分析PIN7的表达分布情况。
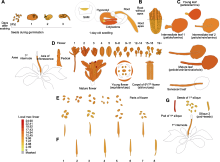
为探明AtFTCD-L的功能, 首先对其在拟南芥各组织中的表达量进行分析。TAIR网站检索结果显示, 该基因在拟南芥的各个发育时期和多个组织器官中均有表达, 尤其在花、叶和根尖中表达量较高(图1A-E), 表明AtFTCD-L在拟南芥不同发育时期的不同组织器官中组成型表达。
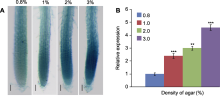
基于组织表达模式分析, AtFTCD-L基因在花、叶与根尖中表达量较高(图1A-E)。为阐明该基因在根中的作用, 我们将GUS-AtFTCD-L种子点播在1/2MS培养基表面, 以及模拟高土壤紧实度生长环境的梯度琼脂培养基中, 对比拟南芥根尖在无阻力与梯度阻力条件下AtFTCD-L的表达情况。结果表明, 随着模拟根生长环境阻力的增加, 拟南芥根尖中GUS染色逐渐加深, 即AtFTCD-L的表达量逐渐增高(图2A); 实时荧光定量PCR检测结果显示, 根尖中AtFTCD-L的表达量随着生长阻力的增大亦不断升高(图2B)。以上实验结果表明, AtFTCD-L可能参与根响应高紧实度阻力。
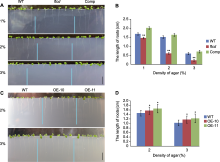
为了确定AtFTCD-L基因在模拟阻力条件下的作用, 我们将野生型、突变体、回复突变体以及过表达材料分别点播到模拟高阻力培养基中, 观察根尖的生长情况。在琼脂浓度为1%的培养基(1%培养基)中, 突变体的根长短于野生型20%; 在2%培养基中, 突变体的根长短于野生型45%; 在3%培养基中, 突变体的根长短于野生型55% (图3A, B), 即根尖在生长阻力越大的环境中生长速度越缓慢, 尤其是突变体植株对高紧实度条件更加敏感, 表明该基因参与响应紧实度阻力。在2%培养基中, 过表达材料的根长较野生型长4%; 在3%培养基中, 过表达材料的根长较野生型增加10% (图3C, D), 表明两种过表达材料的根系均长于野生型根系。综上表明, AtFTCD-L通过响应高紧实度阻力参与拟南芥根的生长。
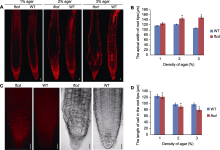
为明确AtFTCD-L基因是否在细胞水平上响应土壤紧实度胁迫, 我们测量了突变体与野生型根尖横向长度与伸长区细胞纵向长度。以生长在梯度培养基中根尖最宽的部位为横向长度, 以伸长区连续5个细胞纵向的长度值除以5代表1个细胞的平均纵向长度。FM4- 64染色观察发现, 高阻力环境下突变体根尖的横向直径略长于野生型植株(图4A)。统计结果显示, 在1%培养基中生长时, 突变体比野生型根尖横向宽6%左右; 在2%培养基中生长时, 突变体比野生型根尖横向宽15%左右; 在3%培养基中生长时, 突变体比野生型根尖横向宽36%左右(图4B)。此外, 在1%培养基中生长时, 突变体比野生型根尖细胞短2%; 在2%培养基中生长时, 突变体比野生型根尖细胞短6%左右; 在3%培养基中生长时, 突变体比野生型根尖细胞短15%左右(图4D)。上述结果表明, 随着模拟阻力逐渐增大, 突变体根尖横向宽度不断加大而纵向长度不断变短。
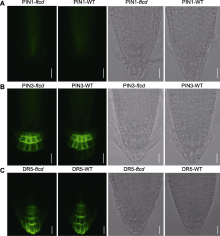
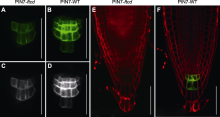
在前期进行荧光漂白恢复实验时, 发现缺失AtFTCD- L会影响细胞中质膜标记蛋白RLK的分泌。为了分析定位于质膜的PIN蛋白是否受AtFTCD-L基因缺失影响表达进而参与调节根系生长, 我们用生长素转运体的内源启动子融合GFP的PIN1、PIN2、PIN3、PIN7和DR5与野生型、突变体与回复突变体种子进行杂交。收集带有GFP标签的稳定表达种子, 点播到2%紧实度阻力培养基中, 以PIN7为代表观察表型, 发现稳定表达PIN7-GFP标签的突变体根长在模拟高紧实度阻力生长条件下同样短于野生型与回复突变体材料(图5)。
本文的其它图/表
|
 首页
首页