植物环核苷酸门控离子通道研究进展
石雅琦, 刘海双, 柯瑾, 马清, 王锁民
植物学报
2025, 60 ( 2):
294-306.
DOI: 10.11983/CBB24119
环核苷酸门控通道(cyclic nucleotide-gated channels, CNGCs)是植物体内重要的阳离子通道, 在调控植物生长发育以及应对冷、热、盐和病原菌等胁迫中发挥重要作用。该文简要概述了植物CNGCs的分类、结构及表达定位, 并重点对其离子选择特征、调控机制以及生物学功能的最新研究进展进行综述, 以期增进对植物CNGCs蛋白的全面认识, 并为后续深入研究提供参考。
| 蛋白 | 亚细胞定位 | 组织特异性 | 可渗透离子 | 功能 | 参考文献 | | AtCNGC1 | 质膜 | 根 | Ca2+、Mg2+和Pb2+ | 根的向地性生长; 重金属离子吸收 | Sunkar et al., 2000; Ma et al., 2006, 2024; Moon et al., 2019 | | AtCNGC2 | 质膜 | 叶脉末端周围的细胞 | Ca2+、Mg2+、K+、Li+、Cs+、Rb+、Sr2+和Ba2+ | 植物免疫; 叶片衰老; 耐热、耐寒性; 成花转变; 生长素稳态 | Leng et al., 1999, 2002; Ma et al., 2010; Finka et al., 2012; Chin et al., 2013; Tian et al., 2019; Cui et al., 2020; Chakraborty et al., 2021 | | AtCNGC3 | 质膜 | 根表皮及皮层、叶脉 | Na+和K+ | 离子稳态; 种子萌发; 植物免疫 | Gobert et al., 2006 | | AtCNGC4 | 质膜 | 叶表皮、胚轴、根毛区、根的伸长区和分生区 | Ca2+、Mg2+、Ba2+、Sr2+、K+ 、Na+和Cs+ | 植物免疫; 程序性细胞死亡; 耐热、耐寒性; 成花转变 | Balagué et al., 2003; Finka et al., 2012; Chin et al., 2013; Tian et al., 2019; Cui et al., 2020 | | AtCNGC5 | 质膜 | 保卫细胞和根毛 | Ca2+、Mg2+和Ba2+ | 耐盐性; 根毛生长; 生长素信号 | Wang et al., 2013; Tan et al., 2020, 2023 | | AtCNGC6 | 质膜 | 保卫细胞和根毛 | Ca2+、Mg2+和Ba2+ | 耐盐性; 根毛生长; 生长素信号; 耐热性 | Gao et al., 2012; Wang et al., 2013; Brost et al., 2019; Tan et al., 2020, 2023 | AtCNGC7
AtCNGC8 | 质膜 | 花粉管尖端 | Ca2+ | 花粉管生长 | Tunc-Ozdemir et al., 2013a; Pan et al., 2019 | | AtCNGC9 | 质膜 | 保卫细胞和根毛 | Ca2+ | 根毛生长; 生长素信号 | Brost et al., 2019; Tan et al., 2020; 2023 | | AtCNGC10 | 质膜、内质网及高尔基体囊泡 | 叶肉细胞、叶片栅栏薄壁组织、叶表皮、根表皮及内皮层和花 | Na+、K+和Pb2+ | 离子稳态; 耐盐性; 根向地性生长; 花粉发育 | Li et al., 2005; Christopher et al., 2007; Guo et al., 2008; Jin et al., 2015; Moon et al., 2019 | | AtCNGC11 | 质膜 | - | Ca2+、Pb2+和Cd2+ | 植物免疫; 细胞死亡 | Urquhart et al., 2007; Moeder et al., 2011; Moon et al., 2019 | | AtCNGC12 | 质膜 | 保卫细胞 | Ca2+和Mg2+ | 植物免疫; 细胞死亡; 生长素信号 | Urquhart et al., 2007; Moeder et al., 2011; DeFalco et al., 2016; Tan et al., 2023 | | AtCNGC13 | - | - | Pb2+和Cd2+ | 重金属离子吸收 | Moon et al., 2019 | | AtCNGC14 | 质膜 | 根毛 | Ca2+ | 根毛生长; 根的向地性
生长 | Shih et al., 2015; Zhang et al., 2017; Brost et al., 2019; Serre et al., 2023 | | AtCNGC15 | 质膜及核膜 | 主根和侧根的根尖、侧根原基和保卫细胞 | Ca2+、Pb2+和Cd2+ | 根的发育 | Moon et al., 2019; Leitão et al., 2019; Wang et al., 2021b | | AtCNGC16 | - | 花粉管 | Ca2+和Cd2+ | 逆境胁迫下的花粉育性 | Tunc-Ozdemir et al., 2013b; Moon et al., 2019 | | AtCNGC17 | 质膜 | - | Ca2+和K+ | 细胞扩张; 耐盐性 | Ladwig et al., 2015 | | AtCNGC18 | 质膜 | 花粉管尖端 | Ca2+ | 花粉管的生长和导向 | Gao et al., 2016; Pan et al., 2019 | | AtCNGC19 | 质膜 | 叶脉周围的叶肉细胞和根韧皮部 | Ca2+、Na+、Pb2+和Cd2+ | 植物免疫; 耐盐性; 植食反应; 细胞死亡 | Kugler et al., 2009; Moon et al., 2019; Meena et al., 2019; Yu et al., 2019, 2023; Jogawat et al., 2020 | | AtCNGC20 | 质膜 | 叶脉周围的叶肉细胞 | Ca2+、Na+和Cd2+ | 植物免疫; 耐盐性; 细胞死亡; 耐寒性 | Kugler et al., 2009; Yu et al., 2019, 2023; Oranab et al., 2021; Peng et al., 2024 |
View table in article
表2
拟南芥环核苷酸门控通道(CNGCs)的定位、离子选择性及其功能
正文中引用本图/表的段落
离子通道是植物体内介导离子沿电化学势梯度跨膜运输的孔道, 直接影响和调控细胞内离子的稳态平衡, 进而在渗透调节、信号转导及养分吸收和长距离运输中发挥关键作用(Pantoja, 2021)。近年来, 多种植物离子通道被鉴定和表征。环核苷酸门控通道(cyclic nucleotide-gated channels, CNGCs)是一种广泛存在于动植物中的阳离子通道, 介导K+、Na+和Ca2+等一价和二价阳离子的转运(Duszyn et al., 2019; Jarratt-Barnham et al., 2021)。CNGCs在动物中的研究较为深入, 在植物中的研究则起步较晚。Schuurink等(1998)在筛选大麦(Hordeum vulgare)钙调素结合转运蛋白(CaM-binding transporter, HvCBT1)的过程中首次克隆到植物CNGC, 随后在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、小麦(Triticum aestivum)和棉花(Gossypium hirsutum)等植物中相继鉴定到其同源基因(K?hler and Neuhaus, 2000; Nawaz et al., 2014; Guo et al., 2018; Lu et al., 2022b)。其中, 模式植物拟南芥CNGCs的研究相对较为系统全面。AtCNGCs家族由20个同源蛋白组成(M?ser et al., 2001), 目前大部分蛋白的离子选择性已在细胞水平上被阐明。随着研究的不断深入, CNGCs在植物生长发育调控、生物及非生物胁迫响应中的重要作用已引起学术界的广泛关注。本文就植物CNGCs的分类、结构、表达定位、离子转运特征、调控机制以及生物学功能进行综述, 旨在为植物CNGCs的深入研究奠定基础。
植物CNGCs成员多数定位于质膜。近年来, 定位于线粒体内膜、叶绿体类囊体膜、核膜、内质网以及高尔基体囊泡的CNGCs相继被鉴定(Duszyn et al., 2019)。在拟南芥中, AtCNGC1、AtCNGC2和AtCNGC3等18个CNGCs定位于质膜(Ma et al., 2006; Chin et al., 2013; Pan et al., 2019)。值得注意的是, 尽管之前的研究认为AtCNGC7、AtCNGC8、AtCNGC19和AtCNGC20定位于液泡膜(Chang et al., 2007; Yuen and Christopher, 2013), 但最新研究表明上述蛋白均定位于质膜而不是液泡膜(Meena et al., 2019; Pan et al., 2019; Yu et al., 2019, 2023; Peng et al., 2024) (表2)。除质膜外, AtCNGC10还定位于内质网及高尔基体囊泡; AtCNGC15及其在蒺藜苜蓿(Medicago truncatula)中的同源蛋白MtCNGC15a/b/c亦定位于核膜(Charpentier et al., 2016; Leit?o et al., 2019)。在水稻中, 根据PSORT亚细胞定位预测, 发现13个OsCNGCs同源蛋白定位于质膜, 其余3个成员(OsCNGC3、OsCNGC7和OsCNGC11)则定位于叶绿体类囊体膜和线粒体内膜(Nawaz et al., 2014)。目前, OsCNGC4、OsCNGC5、OsCNGC8、OsCN- GC9、OsCNGC13、OsCNGC14和OsCNGC16, 已通过在烟草(Nicotiana tabacum)中的瞬时表达证实定位于质膜(Xu et al., 2017; Wang et al., 2019; Cui et al., 2020; Lee et al., 2022)。
作为一类重要的阳离子通道, 多数CNGCs家族蛋白能够介导Ca2+转运, 部分成员对K+、Na+等一价和其它二价阳离子亦具有通透性。在拟南芥20个CNGCs中, 除AtCNGC3、AtCNGC10和AtCNGC13外, 其它17个CNGCs成员均介导Ca2+转运(表2)。目前, 已在非洲爪蟾卵母细胞(Xenopus oocytes)或人体胚胎肾细胞(human embryonic kidney cell, HEK293T)异源系统中通过膜片钳技术证实, AtCNGC1具有Ca2+和Mg2+通透性(Ma et al., 2024)。AtCNGC2和AtCNGC4能够介导环核苷酸(cNMP)依赖的Ca2+和K+吸收, 同时AtCNGC2还可转运Li+、Cs+、Ba2+和Sr2+等一价和二价阳离子(Leng et al., 1999, 2002; Finka et al., 2012); AtCNGC4还具有Na+和Cs+通透性(Balagué et al., 2003)。有趣的是, 当AtCNGC2和AtCNGC4形成复合体时, 对Ca2+、Sr2+、Ba2+和K+的通透性则不依赖于cNMP (Tian et al., 2019)。此外, AtCNGC5、AtCNGC6、AtCNGC7、AtCNGC8、AtCNGC9、AtCNGC14、AtCNGC18、AtCNGC19和AtCNGC20均在爪蟾卵母细胞或HEK293T细胞异源表达系统中具有Ca2+通透性(Pan et al., 2019; Yu et al., 2019; Tan et al., 2020)。在酵母异源表达系统中的分析显示, AtCNGC3具有K+和Na+转运活性, 但不介导Ca2+的转运(Gobert et al., 2006)。将AtCNGC11和12单独或共同转入Ca2+吸收缺陷型酵母菌株中均可促进Ca2+的吸收(Urquhart et al., 2007)。AtCNGC10在大肠杆菌(Escherichia coli)和酵母异源表达系统中表达均可介导K+以及Na+的转运(Li et al., 2005)。本课题组利用酵母及HEK293T异源表达系统分析表明, 霸王的2个ZxCNGC1同源蛋白ZxCNGC1;1以及ZxCNGC1;2均可介导Na+、Ca2+和Mg2+的转运, 但不介导K+的转运(Ma et al., 2024)。另外, 仅在植株水平上分析了部分AtCNGCs成员的离子选择性。例如, Moon等(2019)研究发现AtCNGC1、AtCNGC10或AtCNGC13的单突株系对Pb2+胁迫具有更强的耐受性, 推测上述蛋白介导Pb2+的转运。
有研究表明, 部分CNGC能够形成异源四聚体通道, 通过相互作用介导Ca2+转运, 进而调控植株生长发育及胁迫响应。AtCNGC2和AtCNGC4能够形成Ca2+通道复合体(Tian et al., 2019); AtCNGC7或AtCNGC8能够与AtCNGC18形成异源四聚体, 介导Ca2+向胞质中转运(Pan et al., 2019); AtCNGC19与AtCNGC20也能够形成Ca2+通道复合体(Yu et al., 2019)。此外, 在HEK293T细胞中, ZxCNGC2;1或ZxCNGC4均能介导Ca2+和Mg2+的转运, 但未检测到Na+或K+的转运活性; 而ZxCNGC2;1-ZxCNGC4复合体不仅能介导Ca2+和Mg2+转运, 还能介导Na+转运, 但未检测到K+转运活性(陈芹芹, 2022)。
早期研究认为植物CNGC由cNMP和Ca2+/CaM竞争性调节, CNGC通过与cNMP结合而被激活, 引起Ca2+内流, 使得胞内Ca2+增加, 从而激活Ca2+依赖蛋白激酶(CDPKs), 进而磷酸化下游靶蛋白(Talke et al., 2003)。近年来的研究表明, CNGC的通道活性并非绝对依赖于cNMP的激活。例如, AtCNGC11、AtCNGC12和AtCNGC14的离子通道活性均与cNMP无关(Zhang et al., 2019; Zeb et al., 2020); 在非洲爪蟾卵母细胞异源表达系统中, 多数CNGCs异源四聚体表现出超极化激活的Ca2+通道活性, 不需要cNMP的激活(Tian et al., 2019)。因此, cNMP可能仅是CNGC发挥功能的辅助因子, 而不是必需的门控配体。此外, 胞内增加的Ca2+亦能够激活CaM, 后者通过与CNGC的CaMBDs结合, 抑制cNMP与CNGC的结合, 使得其通道活性减弱, 从而限制Ca2+向胞内转运, 以确保植株对胞内Ca2+信号的精密调节(Hua et al., 2003b; Lemtiri-Chlieh and Berkowitz, 2004)。进一步研究显示, 除CaMBDs外, CaM还能够通过与IQ基序结合进而调控CNGCs (Zhang et al., 2019; Dietrich et al., 2020; Zhao et al., 2021), 表明CaM对植物CNGC的调节较为复杂, 并非完全通过影响cNMP与CNGCs的结合。此外, 有研究表明CNGCs的活性受磷酸化调控。例如, Ca2+依赖的蛋白激酶AtCPK32能够显著增强AtCNGC18的通道活性(Zhou et al., 2014); 胞质激酶BIK1/PBL1能够磷酸化AtCNGC4, 随后激活AtCNGC2-AtCNGC4复合体通道(Tian et al., 2019); 在水稻中, 胞质受体激酶OsRLCK185通过磷酸化激活OsCNGC9, 进而触发防御反应(Wang et al., 2019)。CNGC还受Ca2+非依赖性蛋白激酶的激活。例如, AtCNGC5、AtCNGC6、AtCNGC9和AtCNGC12四个通道能够被Ca2+非依赖性蛋白激酶AtOST1激活并起始胞内Ca2+信号(Yang et al., 2024); 此外, 植物CNGC还能够通过与其它转运蛋白相互作用进而发挥作用, 如AtCNGC15与硝酸盐转运蛋白AtNRT1.1共同组装成“转运体-钙通道复合体(transceptor-channel complex)”, 该复合体能够感应外界硝酸盐浓度变化, 激活或关闭钙离子通道(Wang et al., 2021b)。
植物CNGCs家族成员众多, 分别在不同生长发育阶段的不同器官中表达, 具有多种生物学功能, 尤其是通过介导Ca2+转运调控胞内Ca2+信号, 进而在植物生长发育以及胁迫响应中发挥重要作用(表2)。
CNGCs除参与植物免疫反应外, 还参与植物对植食性昆虫取食的响应。植物防御食草动物取食由茉莉酸(jasmonic acid, JA)依赖的信号级联介导, AtCNGC19的表达能够被食草昆虫的取食行为诱导, 引起Ca2+向胞质中转运, CaM1、CaM4和CaM7感知Ca2+浓度升高并通过激活下游蛋白促进JA的生物合成, 进而触发植物对虫害的防御反应(Yan et al., 2018; Meena et al., 2019)。
干旱胁迫可显著诱导烟草CNGC1 (NtCNGC1)、NtCNGC6和NtCNGC7等在叶中的表达, 表明NtCNGCs参与烟草对干旱的响应(Nawaz et al., 2019)。干旱胁迫下, 甜橙(Citrus sinensis) CsCNGC1.4、CsCNGC2.1和CsCNGC4.2在叶中的表达显著上调, 而CsCNGC4.6的表达显著下降(Zia et al., 2022)。此外, Gao等(2020)发现外源多巴胺可通过促进CNGC和CAM/CML家族基因的表达激活Ca2+信号通路, 从而提高苹果的抗旱性。本课题组对霸王进行转录组分析, 结果表明9个ZxCNGC1s中有7个主要在根中表达, 并且在干旱或盐处理下均上调表达(Ma et al., 2024)。
部分CNGCs已被证实参与重金属离子的吸收和转运。AtCNGC16或者AtCNGC20的突变均减弱了植株对Cd2+胁迫的耐受性; AtCNGC11、AtCNGC13、AtCNGC15或AtCNGC19的突变降低了拟南芥体内Pb2+和Cd2+积累(表2) (Moon et al., 2019)。在烟草中, 多个NtCNGCs在Cd2+胁迫下上调表达, 可能参与植物对Cd2+胁迫的响应(Nawaz et al., 2019)。此外, 拟南芥AtCNGC5、AtCNGC6以及AtCNGC2-AtCNGC4复合体能够介导Ba2+的转运, 可能在植物应对Ba2+胁迫中发挥作用(表2) (Wang et al., 2013; Tian et al., 2019)。
- 未知 ... A putative calcium-permeable cyclic nucleotide-gated channel, CNGC18, regulates polarized pollen tube growth 1 2007 ... 植物CNGCs成员多数定位于质膜.近年来, 定位于线粒体内膜、叶绿体类囊体膜、核膜、内质网以及高尔基体囊泡的CNGCs相继被鉴定(Duszyn et al., 2019).在拟南芥中, AtCNGC1、AtCNGC2和AtCNGC3等18个CNGCs定位于质膜(Ma et al., 2006; Chin et al., 2013; Pan et al., 2019).值得注意的是, 尽管之前的研究认为AtCNGC7、AtCNGC8、AtCNGC19和AtCNGC20定位于液泡膜(Chang et al., 2007; Yuen and Christopher, 2013), 但最新研究表明上述蛋白均定位于质膜而不是液泡膜(Meena et al., 2019; Pan et al., 2019; Yu et al., 2019, 2023; Peng et al., 2024) (表2).除质膜外, AtCNGC10还定位于内质网及高尔基体囊泡; AtCNGC15及其在蒺藜苜蓿(Medicago truncatula)中的同源蛋白MtCNGC15a/b/c亦定位于核膜(Charpentier et al., 2016; Leit?o et al., 2019).在水稻中, 根据PSORT亚细胞定位预测, 发现13个OsCNGCs同源蛋白定位于质膜, 其余3个成员(OsCNGC3、OsCNGC7和OsCNGC11)则定位于叶绿体类囊体膜和线粒体内膜(Nawaz et al., 2014).目前, OsCNGC4、OsCNGC5、OsCNGC8、OsCN- GC9、OsCNGC13、OsCNGC14和OsCNGC16, 已通过在烟草(Nicotiana tabacum)中的瞬时表达证实定位于质膜(Xu et al., 2017; Wang et al., 2019; Cui et al., 2020; Lee et al., 2022). ... Nuclear-localized cyclic nucleotide-gated channels mediate symbiotic calcium oscillations 2 2016 ... 植物CNGCs成员多数定位于质膜.近年来, 定位于线粒体内膜、叶绿体类囊体膜、核膜、内质网以及高尔基体囊泡的CNGCs相继被鉴定(Duszyn et al., 2019).在拟南芥中, AtCNGC1、AtCNGC2和AtCNGC3等18个CNGCs定位于质膜(Ma et al., 2006; Chin et al., 2013; Pan et al., 2019).值得注意的是, 尽管之前的研究认为AtCNGC7、AtCNGC8、AtCNGC19和AtCNGC20定位于液泡膜(Chang et al., 2007; Yuen and Christopher, 2013), 但最新研究表明上述蛋白均定位于质膜而不是液泡膜(Meena et al., 2019; Pan et al., 2019; Yu et al., 2019, 2023; Peng et al., 2024) (表2).除质膜外, AtCNGC10还定位于内质网及高尔基体囊泡; AtCNGC15及其在蒺藜苜蓿(Medicago truncatula)中的同源蛋白MtCNGC15a/b/c亦定位于核膜(Charpentier et al., 2016; Leit?o et al., 2019).在水稻中, 根据PSORT亚细胞定位预测, 发现13个OsCNGCs同源蛋白定位于质膜, 其余3个成员(OsCNGC3、OsCNGC7和OsCNGC11)则定位于叶绿体类囊体膜和线粒体内膜(Nawaz et al., 2014).目前, OsCNGC4、OsCNGC5、OsCNGC8、OsCN- GC9、OsCNGC13、OsCNGC14和OsCNGC16, 已通过在烟草(Nicotiana tabacum)中的瞬时表达证实定位于质膜(Xu et al., 2017; Wang et al., 2019; Cui et al., 2020; Lee et al., 2022). ...
- 未知 ... CYCLIC NUCLEOTIDE-GATED ION CHANNELs 14 and 16 promote tolerance to heat and chilling in rice 5 2020 ... 植物CNGCs成员多数定位于质膜.近年来, 定位于线粒体内膜、叶绿体类囊体膜、核膜、内质网以及高尔基体囊泡的CNGCs相继被鉴定(Duszyn et al., 2019).在拟南芥中, AtCNGC1、AtCNGC2和AtCNGC3等18个CNGCs定位于质膜(Ma et al., 2006; Chin et al., 2013; Pan et al., 2019).值得注意的是, 尽管之前的研究认为AtCNGC7、AtCNGC8、AtCNGC19和AtCNGC20定位于液泡膜(Chang et al., 2007; Yuen and Christopher, 2013), 但最新研究表明上述蛋白均定位于质膜而不是液泡膜(Meena et al., 2019; Pan et al., 2019; Yu et al., 2019, 2023; Peng et al., 2024) (表2).除质膜外, AtCNGC10还定位于内质网及高尔基体囊泡; AtCNGC15及其在蒺藜苜蓿(Medicago truncatula)中的同源蛋白MtCNGC15a/b/c亦定位于核膜(Charpentier et al., 2016; Leit?o et al., 2019).在水稻中, 根据PSORT亚细胞定位预测, 发现13个OsCNGCs同源蛋白定位于质膜, 其余3个成员(OsCNGC3、OsCNGC7和OsCNGC11)则定位于叶绿体类囊体膜和线粒体内膜(Nawaz et al., 2014).目前, OsCNGC4、OsCNGC5、OsCNGC8、OsCN- GC9、OsCNGC13、OsCNGC14和OsCNGC16, 已通过在烟草(Nicotiana tabacum)中的瞬时表达证实定位于质膜(Xu et al., 2017; Wang et al., 2019; Cui et al., 2020; Lee et al., 2022). ...
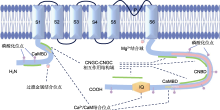
- 未知 ... Stress-induced electrolyte leakage: the role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment 1 2014 ... 植物与动物CNGCs以及植物Shaker型K+电压门控通道的结构高度相似(Talke et al., 2003), 其核心结构为6个跨膜域(S1-S6) (图2).其中, 作为电压传感器的S4带有正电荷, 能够感受和响应膜电位的变化, 具有电压门控的功能(Hua et al., 2003a; 刘海娇等, 2015); 位于S5与S6之间的孔状结构P环(pore loop)具有离子选择性(Chen et al., 2015).植物CNGCs的N端和C端均位于胞质一侧.C端含有1个环核苷酸结合结构域(cyclic nucleotide-binding domain, CNBD)、1个钙调素(calmodulin, CaM)结合结构域(calmodulin-binding domain, CaMBD)以及1个可结合CaM的异亮氨酸-谷氨酰胺(IQ)基序, 其中CNBD和CaMBD部分重叠; C端还包含多个磷酸化位点及CNGC-CNGC相互作用结构域(图2) (Jarratt-Barnham et al., 2021).N端较短, 包含CaMBD、磷酸化位点以及过渡金属结合位点(图2) (Chin et al., 2009; Demidchik et al., 2014).植物CNGCs的CNBD结构域存在一段特异性保守基序“[LI]-X(2)-[GS]- X-[FYIVS]-X-G-X(0,1)-[DE]-LL-X(8,25)-[SA]-X(9)-[VLIT]-E-X-F-X-[IL]”.Zelman等(2012)发现, 基于此基序构建的系统进化树与利用CNGC蛋白全长构建的系统进化树相一致, 因此认为该基序可用于植物CNGCs的鉴定. ... Plant cyclic nucleotide-gated channels: new insights on their functions and regulation 1 2020 ... 早期研究认为植物CNGC由cNMP和Ca2+/CaM竞争性调节, CNGC通过与cNMP结合而被激活, 引起Ca2+内流, 使得胞内Ca2+增加, 从而激活Ca2+依赖蛋白激酶(CDPKs), 进而磷酸化下游靶蛋白(Talke et al., 2003).近年来的研究表明, CNGC的通道活性并非绝对依赖于cNMP的激活.例如, AtCNGC11、AtCNGC12和AtCNGC14的离子通道活性均与cNMP无关(Zhang et al., 2019; Zeb et al., 2020); 在非洲爪蟾卵母细胞异源表达系统中, 多数CNGCs异源四聚体表现出超极化激活的Ca2+通道活性, 不需要cNMP的激活(Tian et al., 2019).因此, cNMP可能仅是CNGC发挥功能的辅助因子, 而不是必需的门控配体.此外, 胞内增加的Ca2+亦能够激活CaM, 后者通过与CNGC的CaMBDs结合, 抑制cNMP与CNGC的结合, 使得其通道活性减弱, 从而限制Ca2+向胞内转运, 以确保植株对胞内Ca2+信号的精密调节(Hua et al., 2003b; Lemtiri-Chlieh and Berkowitz, 2004).进一步研究显示, 除CaMBDs外, CaM还能够通过与IQ基序结合进而调控CNGCs (Zhang et al., 2019; Dietrich et al., 2020; Zhao et al., 2021), 表明CaM对植物CNGC的调节较为复杂, 并非完全通过影响cNMP与CNGCs的结合.此外, 有研究表明CNGCs的活性受磷酸化调控.例如, Ca2+依赖的蛋白激酶AtCPK32能够显著增强AtCNGC18的通道活性(Zhou et al., 2014); 胞质激酶BIK1/PBL1能够磷酸化AtCNGC4, 随后激活AtCNGC2-AtCNGC4复合体通道(Tian et al., 2019); 在水稻中, 胞质受体激酶OsRLCK185通过磷酸化激活OsCNGC9, 进而触发防御反应(Wang et al., 2019).CNGC还受Ca2+非依赖性蛋白激酶的激活.例如, AtCNGC5、AtCNGC6、AtCNGC9和AtCNGC12四个通道能够被Ca2+非依赖性蛋白激酶AtOST1激活并起始胞内Ca2+信号(Yang et al., 2024); 此外, 植物CNGC还能够通过与其它转运蛋白相互作用进而发挥作用, 如AtCNGC15与硝酸盐转运蛋白AtNRT1.1共同组装成“转运体-钙通道复合体(transceptor-channel complex)”, 该复合体能够感应外界硝酸盐浓度变化, 激活或关闭钙离子通道(Wang et al., 2021b). ... Cyclic nucleotide gated channels (CNGCs) in plant signaling—current knowledge and perspectives 2 2019 ... 离子通道是植物体内介导离子沿电化学势梯度跨膜运输的孔道, 直接影响和调控细胞内离子的稳态平衡, 进而在渗透调节、信号转导及养分吸收和长距离运输中发挥关键作用(Pantoja, 2021).近年来, 多种植物离子通道被鉴定和表征.环核苷酸门控通道(cyclic nucleotide-gated channels, CNGCs)是一种广泛存在于动植物中的阳离子通道, 介导K+、Na+和Ca2+等一价和二价阳离子的转运(Duszyn et al., 2019; Jarratt-Barnham et al., 2021).CNGCs在动物中的研究较为深入, 在植物中的研究则起步较晚.Schuurink等(1998)在筛选大麦(Hordeum vulgare)钙调素结合转运蛋白(CaM-binding transporter, HvCBT1)的过程中首次克隆到植物CNGC, 随后在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、小麦(Triticum aestivum)和棉花(Gossypium hirsutum)等植物中相继鉴定到其同源基因(K?hler and Neuhaus, 2000; Nawaz et al., 2014; Guo et al., 2018; Lu et al., 2022b).其中, 模式植物拟南芥CNGCs的研究相对较为系统全面.AtCNGCs家族由20个同源蛋白组成(M?ser et al., 2001), 目前大部分蛋白的离子选择性已在细胞水平上被阐明.随着研究的不断深入, CNGCs在植物生长发育调控、生物及非生物胁迫响应中的重要作用已引起学术界的广泛关注.本文就植物CNGCs的分类、结构、表达定位、离子转运特征、调控机制以及生物学功能进行综述, 旨在为植物CNGCs的深入研究奠定基础. ...
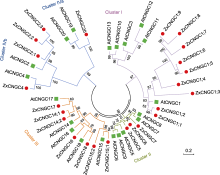
- 未知 ... Identification and analysis of CNGC family genes in potato (Solanum tuberosum) 1 2023 ... 根据氨基酸序列构建的系统进化树, 可将AtCNGCs分为4个亚家族(I、II、III和IV).其中, 第I、II、III亚家族之间的亲缘关系较近; 第IV亚家族与其它亚家族的关系则相对较远, 并包括IVa和IVb两个亚组(Talke et al., 2003).研究发现, 小麦、水稻、大豆(Glycine max)、白菜(Brassica rapa)、马铃薯(Solanum tuberosum)以及4个棉属植物CNGCs家族的系统发育关系均符合上述分类(Nawaz et al., 2014; Guo et al., 2018; Baloch et al., 2021; Lu et al., 2022b; Cui et al., 2023; 郝宇瑞等, 2023).本课题组对荒漠植物霸王(Zygophyllum xanthoxylum)进行全基因组测序, 鉴定到24个ZxCNGCs成员, 同样分为I、II、III、IV四个亚家族, 并且第IV亚族包含IVa和IVb两个亚组(图1); 有趣的是, 其中ZxCNGC1s有9个同源基因, 与其近缘物种四合木(Tetraena mongolica)、盐生植物小花碱茅(Puccinellia tenuiflorav)和藜麦(Chenopodium quinoa)以及甜土植物桃金娘(Rhodomyrtus tomentosa)、毛果杨(Populus trichocarpa)、葡萄(Vitis vinifera)和拟南芥相比发生了明显扩张(Ma et al., 2024) (表1). ... 马铃薯CNGC家族全基因组鉴定和组织特异性分析 1 2023 ... 根据氨基酸序列构建的系统进化树, 可将AtCNGCs分为4个亚家族(I、II、III和IV).其中, 第I、II、III亚家族之间的亲缘关系较近; 第IV亚家族与其它亚家族的关系则相对较远, 并包括IVa和IVb两个亚组(Talke et al., 2003).研究发现, 小麦、水稻、大豆(Glycine max)、白菜(Brassica rapa)、马铃薯(Solanum tuberosum)以及4个棉属植物CNGCs家族的系统发育关系均符合上述分类(Nawaz et al., 2014; Guo et al., 2018; Baloch et al., 2021; Lu et al., 2022b; Cui et al., 2023; 郝宇瑞等, 2023).本课题组对荒漠植物霸王(Zygophyllum xanthoxylum)进行全基因组测序, 鉴定到24个ZxCNGCs成员, 同样分为I、II、III、IV四个亚家族, 并且第IV亚族包含IVa和IVb两个亚组(图1); 有趣的是, 其中ZxCNGC1s有9个同源基因, 与其近缘物种四合木(Tetraena mongolica)、盐生植物小花碱茅(Puccinellia tenuiflorav)和藜麦(Chenopodium quinoa)以及甜土植物桃金娘(Rhodomyrtus tomentosa)、毛果杨(Populus trichocarpa)、葡萄(Vitis vinifera)和拟南芥相比发生了明显扩张(Ma et al., 2024) (表1). ... Plants do it differently. A new basis for potassium/sodium selectivity in the pore of an ion channel 1 2003a ... 植物与动物CNGCs以及植物Shaker型K+电压门控通道的结构高度相似(Talke et al., 2003), 其核心结构为6个跨膜域(S1-S6) (图2).其中, 作为电压传感器的S4带有正电荷, 能够感受和响应膜电位的变化, 具有电压门控的功能(Hua et al., 2003a; 刘海娇等, 2015); 位于S5与S6之间的孔状结构P环(pore loop)具有离子选择性(Chen et al., 2015).植物CNGCs的N端和C端均位于胞质一侧.C端含有1个环核苷酸结合结构域(cyclic nucleotide-binding domain, CNBD)、1个钙调素(calmodulin, CaM)结合结构域(calmodulin-binding domain, CaMBD)以及1个可结合CaM的异亮氨酸-谷氨酰胺(IQ)基序, 其中CNBD和CaMBD部分重叠; C端还包含多个磷酸化位点及CNGC-CNGC相互作用结构域(图2) (Jarratt-Barnham et al., 2021).N端较短, 包含CaMBD、磷酸化位点以及过渡金属结合位点(图2) (Chin et al., 2009; Demidchik et al., 2014).植物CNGCs的CNBD结构域存在一段特异性保守基序“[LI]-X(2)-[GS]- X-[FYIVS]-X-G-X(0,1)-[DE]-LL-X(8,25)-[SA]-X(9)-[VLIT]-E-X-F-X-[IL]”.Zelman等(2012)发现, 基于此基序构建的系统进化树与利用CNGC蛋白全长构建的系统进化树相一致, 因此认为该基序可用于植物CNGCs的鉴定. ... Functional interaction of calmodulin with a plant cyclic nucleotide gated cation channel 1 2003b ... 早期研究认为植物CNGC由cNMP和Ca2+/CaM竞争性调节, CNGC通过与cNMP结合而被激活, 引起Ca2+内流, 使得胞内Ca2+增加, 从而激活Ca2+依赖蛋白激酶(CDPKs), 进而磷酸化下游靶蛋白(Talke et al., 2003).近年来的研究表明, CNGC的通道活性并非绝对依赖于cNMP的激活.例如, AtCNGC11、AtCNGC12和AtCNGC14的离子通道活性均与cNMP无关(Zhang et al., 2019; Zeb et al., 2020); 在非洲爪蟾卵母细胞异源表达系统中, 多数CNGCs异源四聚体表现出超极化激活的Ca2+通道活性, 不需要cNMP的激活(Tian et al., 2019).因此, cNMP可能仅是CNGC发挥功能的辅助因子, 而不是必需的门控配体.此外, 胞内增加的Ca2+亦能够激活CaM, 后者通过与CNGC的CaMBDs结合, 抑制cNMP与CNGC的结合, 使得其通道活性减弱, 从而限制Ca2+向胞内转运, 以确保植株对胞内Ca2+信号的精密调节(Hua et al., 2003b; Lemtiri-Chlieh and Berkowitz, 2004).进一步研究显示, 除CaMBDs外, CaM还能够通过与IQ基序结合进而调控CNGCs (Zhang et al., 2019; Dietrich et al., 2020; Zhao et al., 2021), 表明CaM对植物CNGC的调节较为复杂, 并非完全通过影响cNMP与CNGCs的结合.此外, 有研究表明CNGCs的活性受磷酸化调控.例如, Ca2+依赖的蛋白激酶AtCPK32能够显著增强AtCNGC18的通道活性(Zhou et al., 2014); 胞质激酶BIK1/PBL1能够磷酸化AtCNGC4, 随后激活AtCNGC2-AtCNGC4复合体通道(Tian et al., 2019); 在水稻中, 胞质受体激酶OsRLCK185通过磷酸化激活OsCNGC9, 进而触发防御反应(Wang et al., 2019).CNGC还受Ca2+非依赖性蛋白激酶的激活.例如, AtCNGC5、AtCNGC6、AtCNGC9和AtCNGC12四个通道能够被Ca2+非依赖性蛋白激酶AtOST1激活并起始胞内Ca2+信号(Yang et al., 2024); 此外, 植物CNGC还能够通过与其它转运蛋白相互作用进而发挥作用, 如AtCNGC15与硝酸盐转运蛋白AtNRT1.1共同组装成“转运体-钙通道复合体(transceptor-channel complex)”, 该复合体能够感应外界硝酸盐浓度变化, 激活或关闭钙离子通道(Wang et al., 2021b). ... Harnessing salt for woody biomass production 1 2012 ... 在拟南芥中, AtCNGC1、AtCNGC10、AtCNGC19和AtCNGC20的表达丰度受盐胁迫诱导(Balagué et al., 2003; Jin et al., 2015).在NaCl或KCl处理下, atcngc3突变体幼苗的长势略优于野生型, 表明AtCNGC3负调控植物的耐盐性(Gobert et al., 2006).盐胁迫下AtCNGC19和AtCNGC20在根中的表达量升高, 且超表达AtCNGC19或AtCNGC20能够提高植株的耐盐性(Oranab et al., 2021).BrCNGC7、BrCNGC10、BrCNGC12、BrCNGC13、BrCNGC19、BrCNGC22和BrCNGC23在白菜根中的表达受到盐胁迫诱导上调(Li et al., 2019).在棉花中, GhCNGC32或者GhCNGC35的沉默均会降低植株的耐盐性(Lu et al., 2022b).此外, 本课题组在分析荒漠植物抗逆机制的过程中, 发现多浆旱生植物霸王生境土壤中有效Na+含量仅为盐生植物盐地碱蓬(Suaeda salsa)的1/66, 但其叶中能够积累与盐地碱蓬相当的Na+, 并将其区域化至液泡中用于渗透调节, 表明霸王根系具有极强的Na+吸收能力(Wang et al., 2002, 2004; Wu et al., 2011; Janz and Polle, 2012; Ma et al., 2012; Yuan et al., 2015).最近的研究表明, 霸王ZxCNGC1;1和ZxCNGC1;2对Na+具有极高的通透性, 在正常的胞内电压(-80 - -120 mV)下即可开放; 而甜土植物水稻OsCNGC1仅在消耗过多能量使胞内电压达到-180 mV时才能开放; 模式植物拟南芥AtCNGC1则在任何检测电压下均无Na+通透性(Ma et al., 2024).可见, ZxCNGC1;1和ZxCNGC1;2能够介导“节能型”的Na+吸收, 进而在维持旱生植物霸王的积盐特性中发挥重要作用, 这揭示了积盐型植物高效吸收Na+的分子基础(Ma et al., 2024). ... The complex story of plant cyclic nucleotide-gated channels 2 2021 ... 离子通道是植物体内介导离子沿电化学势梯度跨膜运输的孔道, 直接影响和调控细胞内离子的稳态平衡, 进而在渗透调节、信号转导及养分吸收和长距离运输中发挥关键作用(Pantoja, 2021).近年来, 多种植物离子通道被鉴定和表征.环核苷酸门控通道(cyclic nucleotide-gated channels, CNGCs)是一种广泛存在于动植物中的阳离子通道, 介导K+、Na+和Ca2+等一价和二价阳离子的转运(Duszyn et al., 2019; Jarratt-Barnham et al., 2021).CNGCs在动物中的研究较为深入, 在植物中的研究则起步较晚.Schuurink等(1998)在筛选大麦(Hordeum vulgare)钙调素结合转运蛋白(CaM-binding transporter, HvCBT1)的过程中首次克隆到植物CNGC, 随后在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、小麦(Triticum aestivum)和棉花(Gossypium hirsutum)等植物中相继鉴定到其同源基因(K?hler and Neuhaus, 2000; Nawaz et al., 2014; Guo et al., 2018; Lu et al., 2022b).其中, 模式植物拟南芥CNGCs的研究相对较为系统全面.AtCNGCs家族由20个同源蛋白组成(M?ser et al., 2001), 目前大部分蛋白的离子选择性已在细胞水平上被阐明.随着研究的不断深入, CNGCs在植物生长发育调控、生物及非生物胁迫响应中的重要作用已引起学术界的广泛关注.本文就植物CNGCs的分类、结构、表达定位、离子转运特征、调控机制以及生物学功能进行综述, 旨在为植物CNGCs的深入研究奠定基础. ...
- 未知 ... Genome-wide analysis of cyclic nucleotide-gated channel genes related to pollen development in rice 1 2022 ... 植物CNGCs成员多数定位于质膜.近年来, 定位于线粒体内膜、叶绿体类囊体膜、核膜、内质网以及高尔基体囊泡的CNGCs相继被鉴定(Duszyn et al., 2019).在拟南芥中, AtCNGC1、AtCNGC2和AtCNGC3等18个CNGCs定位于质膜(Ma et al., 2006; Chin et al., 2013; Pan et al., 2019).值得注意的是, 尽管之前的研究认为AtCNGC7、AtCNGC8、AtCNGC19和AtCNGC20定位于液泡膜(Chang et al., 2007; Yuen and Christopher, 2013), 但最新研究表明上述蛋白均定位于质膜而不是液泡膜(Meena et al., 2019; Pan et al., 2019; Yu et al., 2019, 2023; Peng et al., 2024) (表2).除质膜外, AtCNGC10还定位于内质网及高尔基体囊泡; AtCNGC15及其在蒺藜苜蓿(Medicago truncatula)中的同源蛋白MtCNGC15a/b/c亦定位于核膜(Charpentier et al., 2016; Leit?o et al., 2019).在水稻中, 根据PSORT亚细胞定位预测, 发现13个OsCNGCs同源蛋白定位于质膜, 其余3个成员(OsCNGC3、OsCNGC7和OsCNGC11)则定位于叶绿体类囊体膜和线粒体内膜(Nawaz et al., 2014).目前, OsCNGC4、OsCNGC5、OsCNGC8、OsCN- GC9、OsCNGC13、OsCNGC14和OsCNGC16, 已通过在烟草(Nicotiana tabacum)中的瞬时表达证实定位于质膜(Xu et al., 2017; Wang et al., 2019; Cui et al., 2020; Lee et al., 2022). ... Nuclear calcium signatures are associated with root development 2 2019 ... 植物CNGCs成员多数定位于质膜.近年来, 定位于线粒体内膜、叶绿体类囊体膜、核膜、内质网以及高尔基体囊泡的CNGCs相继被鉴定(Duszyn et al., 2019).在拟南芥中, AtCNGC1、AtCNGC2和AtCNGC3等18个CNGCs定位于质膜(Ma et al., 2006; Chin et al., 2013; Pan et al., 2019).值得注意的是, 尽管之前的研究认为AtCNGC7、AtCNGC8、AtCNGC19和AtCNGC20定位于液泡膜(Chang et al., 2007; Yuen and Christopher, 2013), 但最新研究表明上述蛋白均定位于质膜而不是液泡膜(Meena et al., 2019; Pan et al., 2019; Yu et al., 2019, 2023; Peng et al., 2024) (表2).除质膜外, AtCNGC10还定位于内质网及高尔基体囊泡; AtCNGC15及其在蒺藜苜蓿(Medicago truncatula)中的同源蛋白MtCNGC15a/b/c亦定位于核膜(Charpentier et al., 2016; Leit?o et al., 2019).在水稻中, 根据PSORT亚细胞定位预测, 发现13个OsCNGCs同源蛋白定位于质膜, 其余3个成员(OsCNGC3、OsCNGC7和OsCNGC11)则定位于叶绿体类囊体膜和线粒体内膜(Nawaz et al., 2014).目前, OsCNGC4、OsCNGC5、OsCNGC8、OsCN- GC9、OsCNGC13、OsCNGC14和OsCNGC16, 已通过在烟草(Nicotiana tabacum)中的瞬时表达证实定位于质膜(Xu et al., 2017; Wang et al., 2019; Cui et al., 2020; Lee et al., 2022). ...
- 未知 ... Cyclic adenosine monophosphate regulates calcium channels in the plasma membrane of Arabidopsis leaf guard and mesophyll cells 1 2004 ... 早期研究认为植物CNGC由cNMP和Ca2+/CaM竞争性调节, CNGC通过与cNMP结合而被激活, 引起Ca2+内流, 使得胞内Ca2+增加, 从而激活Ca2+依赖蛋白激酶(CDPKs), 进而磷酸化下游靶蛋白(Talke et al., 2003).近年来的研究表明, CNGC的通道活性并非绝对依赖于cNMP的激活.例如, AtCNGC11、AtCNGC12和AtCNGC14的离子通道活性均与cNMP无关(Zhang et al., 2019; Zeb et al., 2020); 在非洲爪蟾卵母细胞异源表达系统中, 多数CNGCs异源四聚体表现出超极化激活的Ca2+通道活性, 不需要cNMP的激活(Tian et al., 2019).因此, cNMP可能仅是CNGC发挥功能的辅助因子, 而不是必需的门控配体.此外, 胞内增加的Ca2+亦能够激活CaM, 后者通过与CNGC的CaMBDs结合, 抑制cNMP与CNGC的结合, 使得其通道活性减弱, 从而限制Ca2+向胞内转运, 以确保植株对胞内Ca2+信号的精密调节(Hua et al., 2003b; Lemtiri-Chlieh and Berkowitz, 2004).进一步研究显示, 除CaMBDs外, CaM还能够通过与IQ基序结合进而调控CNGCs (Zhang et al., 2019; Dietrich et al., 2020; Zhao et al., 2021), 表明CaM对植物CNGC的调节较为复杂, 并非完全通过影响cNMP与CNGCs的结合.此外, 有研究表明CNGCs的活性受磷酸化调控.例如, Ca2+依赖的蛋白激酶AtCPK32能够显著增强AtCNGC18的通道活性(Zhou et al., 2014); 胞质激酶BIK1/PBL1能够磷酸化AtCNGC4, 随后激活AtCNGC2-AtCNGC4复合体通道(Tian et al., 2019); 在水稻中, 胞质受体激酶OsRLCK185通过磷酸化激活OsCNGC9, 进而触发防御反应(Wang et al., 2019).CNGC还受Ca2+非依赖性蛋白激酶的激活.例如, AtCNGC5、AtCNGC6、AtCNGC9和AtCNGC12四个通道能够被Ca2+非依赖性蛋白激酶AtOST1激活并起始胞内Ca2+信号(Yang et al., 2024); 此外, 植物CNGC还能够通过与其它转运蛋白相互作用进而发挥作用, 如AtCNGC15与硝酸盐转运蛋白AtNRT1.1共同组装成“转运体-钙通道复合体(transceptor-channel complex)”, 该复合体能够感应外界硝酸盐浓度变化, 激活或关闭钙离子通道(Wang et al., 2021b). ... Electrophysiological analysis of cloned cyclic nucleotide-gated ion channels 2 2002 ... Localization, ion selectivity and function of cyclic nucleotide-gated channels (CNGCs) in Arabidopsis
- 未知 ... Phylogenetic relationships within cation transporter families of Arabidopsis 1 2001 ... 离子通道是植物体内介导离子沿电化学势梯度跨膜运输的孔道, 直接影响和调控细胞内离子的稳态平衡, 进而在渗透调节、信号转导及养分吸收和长距离运输中发挥关键作用(Pantoja, 2021).近年来, 多种植物离子通道被鉴定和表征.环核苷酸门控通道(cyclic nucleotide-gated channels, CNGCs)是一种广泛存在于动植物中的阳离子通道, 介导K+、Na+和Ca2+等一价和二价阳离子的转运(Duszyn et al., 2019; Jarratt-Barnham et al., 2021).CNGCs在动物中的研究较为深入, 在植物中的研究则起步较晚.Schuurink等(1998)在筛选大麦(Hordeum vulgare)钙调素结合转运蛋白(CaM-binding transporter, HvCBT1)的过程中首次克隆到植物CNGC, 随后在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、小麦(Triticum aestivum)和棉花(Gossypium hirsutum)等植物中相继鉴定到其同源基因(K?hler and Neuhaus, 2000; Nawaz et al., 2014; Guo et al., 2018; Lu et al., 2022b).其中, 模式植物拟南芥CNGCs的研究相对较为系统全面.AtCNGCs家族由20个同源蛋白组成(M?ser et al., 2001), 目前大部分蛋白的离子选择性已在细胞水平上被阐明.随着研究的不断深入, CNGCs在植物生长发育调控、生物及非生物胁迫响应中的重要作用已引起学术界的广泛关注.本文就植物CNGCs的分类、结构、表达定位、离子转运特征、调控机制以及生物学功能进行综述, 旨在为植物CNGCs的深入研究奠定基础. ... The Ca2+ channel CNGC19 regulates Arabidopsis defense against spodoptera herbivory 3 2019 ... 植物CNGCs成员多数定位于质膜.近年来, 定位于线粒体内膜、叶绿体类囊体膜、核膜、内质网以及高尔基体囊泡的CNGCs相继被鉴定(Duszyn et al., 2019).在拟南芥中, AtCNGC1、AtCNGC2和AtCNGC3等18个CNGCs定位于质膜(Ma et al., 2006; Chin et al., 2013; Pan et al., 2019).值得注意的是, 尽管之前的研究认为AtCNGC7、AtCNGC8、AtCNGC19和AtCNGC20定位于液泡膜(Chang et al., 2007; Yuen and Christopher, 2013), 但最新研究表明上述蛋白均定位于质膜而不是液泡膜(Meena et al., 2019; Pan et al., 2019; Yu et al., 2019, 2023; Peng et al., 2024) (表2).除质膜外, AtCNGC10还定位于内质网及高尔基体囊泡; AtCNGC15及其在蒺藜苜蓿(Medicago truncatula)中的同源蛋白MtCNGC15a/b/c亦定位于核膜(Charpentier et al., 2016; Leit?o et al., 2019).在水稻中, 根据PSORT亚细胞定位预测, 发现13个OsCNGCs同源蛋白定位于质膜, 其余3个成员(OsCNGC3、OsCNGC7和OsCNGC11)则定位于叶绿体类囊体膜和线粒体内膜(Nawaz et al., 2014).目前, OsCNGC4、OsCNGC5、OsCNGC8、OsCN- GC9、OsCNGC13、OsCNGC14和OsCNGC16, 已通过在烟草(Nicotiana tabacum)中的瞬时表达证实定位于质膜(Xu et al., 2017; Wang et al., 2019; Cui et al., 2020; Lee et al., 2022). ...
- 未知 ... An apple cyclic nucleotide-gated ion channel gene highly responsive to Botryosphaeria dothidea infection enhances the susceptibility of Nicotiana benthamiana to bacterial and fungal pathogens 1 2018 ... 植物中多个CNGCs成员参与响应Ca2+依赖的免疫反应, 其中对AtCNGC2和AtCNGC4的研究最为深入.Tian等(2019)与Wang等(2024)阐明了AtCNGC2- AtCNGC4复合体在病原体相关分子模式(pathogen- associated molecular patterns, PAMPs)触发的免疫反应(PAMP-triggered immunity, PTI)中介导的Ca2+信号转导途径: 在静息状态下, 植物模式识别受体(pattern-recognition receptors, PRRs)处于无活性状态, AtCNGC2-AtCNGC4复合体被CaMs抑制而关闭; 当病原体入侵时, PAMPs与PRRs结合, 并激活PRR的效应激酶BIK1 (BOTRYTIS-INDUCED KINASE1), 随后, AtCNGC2-AtCNGC4通道被BIK1磷酸化并激活, 从而引发胞质中游离的Ca2+ ([Ca2+]cyt)增加, 此过程起始Ca2+信号并产生免疫反应; 当[Ca2+]cyt升高后, BIK1/PBL1磷酸化并激活液泡膜上的Ca2+/H+逆向转运蛋白(Ca2+/H+ antiporters, CAXs), 将Ca2+区域化至液泡中, 降低胞质中的Ca2+浓度以解除Ca2+信号, 从而调节生长和免疫响应之间的平衡.此外, AtCNGC3、AtCNGC11、AtCNGC12、AtCNGC19和AtCNGC20也被证实是病原体防御反应的正调节因子(Moeder et al., 2011; Yu et al., 2019; Jogawat et al., 2020; 刘梦娇, 2021).水稻中也有多个OsCNGCs家族蛋白参与病原菌胁迫响应, 其中OsCNGC5和OsCNGC9正调控水稻的抗病性(Wang et al., 2019; Zheng et al., 2024).在接种不同病原菌后, 苹果(Malus pumila)、烟草及大豆等植物中大量CNGCs的表达水平发生显著变化, 表明CNGCs可能在响应病原菌胁迫中发挥重要作用(Zhang et al., 2018; Nawaz et al., 2019; Cui et al., 2023). ... Genome-wide analysis and expression of Cyclic Nucleotide-Gated Ion Channel (CNGC) family genes under cold stress in mango (Mangifera indica) 1 2023 ... 植物CNGCs在温度胁迫诱导的Ca2+信号转导中发挥关键作用.部分CNGCs参与响应热胁迫.例如, AtCNGC6介导热胁迫诱导的Ca2+内流, 并影响热激蛋白的表达以增强植株的耐热性(Peng et al., 2019); 白菜CNGC1 (BrCNGC1)、BrCNGC3、BrCNGC10、BrCNGC17和BrCNGC29在高温胁迫下上调表达(Li et al., 2019).此外, CNGCs亦在植物响应冷胁迫中发挥作用.最新研究表明, AtCNGC20通过介导Ca2+的转运正调节拟南芥的耐寒性, 在低温胁迫早期, 酪氨酸硫酸肽受体激酶PSY1R (Plant Peptide Containing Sulfated Tyrosine1 Receptor)被激活, 进而磷酸化AtCNGC20并增强其活性; 而在长期低温胁迫下, 冷响应蛋白激酶CRPK1 (Cold-Responsive Protein Kinase1)磷酸化并降解AtCNGC20, 从而降低植株的耐寒性(Peng et al., 2024).OsCNGC9也可通过OsSAPK8蛋白激酶的磷酸化正调控水稻的耐寒性(Wang et al., 2021a).在低温胁迫下, 甘蓝(B. ole- racea) BoCNGC17和BoCNGC23、烟草NtCNGC6和NtCNGC7、芒果(Mangifera indica) MiCNGC9和MiCNGC13以及茄子(Solanum melongena) SmCNGC1a的表达水平均显著上调(Kakar et al., 2017; Nawaz et al., 2019; Zhang et al., 2023; Jiang et al., 2023).此外, CNGC2和CNGC4在拟南芥耐热性和耐寒性中均发挥重要作用.AtCNGC2和AtCNGC4对植株耐热性的影响依赖于发育阶段, AtCNGC2的缺失导致植物幼苗期耐热性增强, 但生殖期耐热性降低(Finka et al., 2012; Katano et al., 2018; Lu et al., 2022a); 同时, 二者正调控植物幼苗期的耐寒性, 其单突变体株系在低温条件下均表现出下胚轴缩短、鲜重及存活率降低(Cui et al., 2020).水稻OsCNGC14和OsCNGC16也在植株耐热和耐寒中发挥作用(Cui et al., 2020). ... Electrophysiological studies revealed CaM1-mediated regulation of the Arabidopsis calcium channel CNGC12 2 2019 ... 早期研究认为植物CNGC由cNMP和Ca2+/CaM竞争性调节, CNGC通过与cNMP结合而被激活, 引起Ca2+内流, 使得胞内Ca2+增加, 从而激活Ca2+依赖蛋白激酶(CDPKs), 进而磷酸化下游靶蛋白(Talke et al., 2003).近年来的研究表明, CNGC的通道活性并非绝对依赖于cNMP的激活.例如, AtCNGC11、AtCNGC12和AtCNGC14的离子通道活性均与cNMP无关(Zhang et al., 2019; Zeb et al., 2020); 在非洲爪蟾卵母细胞异源表达系统中, 多数CNGCs异源四聚体表现出超极化激活的Ca2+通道活性, 不需要cNMP的激活(Tian et al., 2019).因此, cNMP可能仅是CNGC发挥功能的辅助因子, 而不是必需的门控配体.此外, 胞内增加的Ca2+亦能够激活CaM, 后者通过与CNGC的CaMBDs结合, 抑制cNMP与CNGC的结合, 使得其通道活性减弱, 从而限制Ca2+向胞内转运, 以确保植株对胞内Ca2+信号的精密调节(Hua et al., 2003b; Lemtiri-Chlieh and Berkowitz, 2004).进一步研究显示, 除CaMBDs外, CaM还能够通过与IQ基序结合进而调控CNGCs (Zhang et al., 2019; Dietrich et al., 2020; Zhao et al., 2021), 表明CaM对植物CNGC的调节较为复杂, 并非完全通过影响cNMP与CNGCs的结合.此外, 有研究表明CNGCs的活性受磷酸化调控.例如, Ca2+依赖的蛋白激酶AtCPK32能够显著增强AtCNGC18的通道活性(Zhou et al., 2014); 胞质激酶BIK1/PBL1能够磷酸化AtCNGC4, 随后激活AtCNGC2-AtCNGC4复合体通道(Tian et al., 2019); 在水稻中, 胞质受体激酶OsRLCK185通过磷酸化激活OsCNGC9, 进而触发防御反应(Wang et al., 2019).CNGC还受Ca2+非依赖性蛋白激酶的激活.例如, AtCNGC5、AtCNGC6、AtCNGC9和AtCNGC12四个通道能够被Ca2+非依赖性蛋白激酶AtOST1激活并起始胞内Ca2+信号(Yang et al., 2024); 此外, 植物CNGC还能够通过与其它转运蛋白相互作用进而发挥作用, 如AtCNGC15与硝酸盐转运蛋白AtNRT1.1共同组装成“转运体-钙通道复合体(transceptor-channel complex)”, 该复合体能够感应外界硝酸盐浓度变化, 激活或关闭钙离子通道(Wang et al., 2021b). ...
本文的其它图/表
|

 首页
首页