|
|
|||||||||||||||||||||||||||||
|
质膜蛋白动力学的调控及其研究方法
植物学报
2023, 58 (4):
590-601.
DOI: 10.11983/CBB22102
质膜蛋白是细胞膜的重要组分之一, 在细胞的物质转运、离子交换、信号转导以及代谢过程中起着重要作用。质膜蛋白在膜上是运动的, 生长发育和环境因素均可改变其运动方式。因此, 研究影响质膜蛋白运动的因素及调控机制对于理解植物生长发育和应对环境改变至关重要。近年来, 显微技术发展迅速, 使得关于质膜蛋白动力学调控机制的研究逐步深入。该文详细介绍了质膜蛋白动力学及其影响因素, 概述了近几年在质膜蛋白动力学研究中常用的显微成像技术, 以期为深入研究质膜蛋白的生物学功能提供参考。
表1
质膜蛋白动力学研究方法的比较
正文中引用本图/表的段落
细胞质膜(plasma membrane, PM)是细胞原生质体外围的膜性结构, 对维持细胞内环境的稳定及完成细胞生长发育过程均具有重要作用(邱丽丽等, 2017)。蛋白质作为生命活动的主要载体, 对细胞膜功能的发挥至关重要, 当细胞膜感受到外界刺激或环境变化时, 其上的蛋白通常会迅速发生侧向位移和聚合等运动, 进而将信号传递下去(苏泊丹等, 2016; Jozefowicz et al., 2018)。许多研究表明, 在植物细胞中, 细胞壁、细胞骨架和膜筏等因素均会对PM蛋白的运动产生影响。细胞壁作为植物细胞的特有结构, 可限制PM蛋白的侧向扩散(Martinière et al., 2012), 缩短PM与细胞壁之间的距离使PM蛋白运动减慢(Li et al., 2013)。细胞骨架是一个动态的、三维丝状蛋白网络结构, 可通过对PM进行分区来限制膜蛋白的扩散, 并影响蛋白的寡聚化和信号转导过程(Jaqaman and Grinstein, 2012)。膜筏作为细胞膜上的一种微结构域, 对膜蛋白的侧向扩散也起一定的限制作用。例如, 对定位于植物质膜上的超敏反应(hypersensitive induced reaction, HIR)蛋白的动力学分析表明, AtHIR1的动力学和寡聚化状态等均受膜筏和细胞骨架的调控, 并且膜筏和细胞骨架参与植物免疫相关复合体的形成(Lv et al., 2017)。
质膜蛋白是质膜功能发挥的主要承担者。在植物细胞中, 质膜蛋白的动态行为多与植物的生长发育、代谢、信号转导和抗病等生命活动息息相关。例如, Xue等(2018)应用单分子研究体系, 在高时空分辨率上揭示了蓝光对细胞质膜上的向光素受体动力学调控机制, 即蓝光驱动phot1二聚化, 进而发生自身磷酸化, 促使其在膜上扩散运动加快并发生胞吞现象, 且膜微区的存在为phot1介导的蓝光信号传递提供了重要平台。Zhang等(2019)采用单颗粒示踪技术对NRT1.1蛋白在质膜上的扩散范围和扩散系数进行了分析, 结果发现在低浓度硝酸根环境中, 定位于膜筏微区中的NRT1.1蛋白分子中长距离运动所占比例较高, 很可能对其生长素转运功能有重要促进作用。研究发现, 膜筏标记蛋白Flot1 (Flotillin1)经过MβCD (甲基-β-环糊精, 可破坏膜筏结构)处理后, 动力学特征显著降低, 并且早期flg22信号级联发生改变, 表明微结构域可能是相关信号转导所必需。使用超分辨率显微镜对Flot1进行检查进一步支持了这一观点, 即质膜上蛋白质的动力学与信号转导有关(Rostoks et al., 2003; Son et al., 2014)。因此, 研究质膜蛋白的动力学有利于从分子水平上了解细胞的代谢活动, 对全面解析细胞膜的功能具有重要意义。
质膜蛋白是细胞膜的重要组分, 也是许多生物学功能的执行者。PM蛋白在膜上的排列并不固定, 而是处于动态变化过程中。因此, 在不同组织中, 质膜蛋白在膜上的分布与运动状况可能存在差异。研究人员使用全内反射荧光显微镜(total internal reflection fluorescence microscopy, TIRFM)对拟南芥(Arabidopsis thaliana)质膜上BSK1蛋白和PM标记物LTi6a进行了观察, 结果表明与LTi6aGFP的均匀分布相反, BSK1-GFP在PM上分布不均匀, 且呈现高度动态性(Su et al., 2021)。Xu等(2022)使用TIRFM对拟南芥质膜上的HIR1 (hypersensitive induced reaction 1)蛋白进行观察, 结果表明HIR1蛋白在植物不同组织中的分布不同, 具异质性。
之前的研究表明, 当外部环境改变或存在胞内信号时, 质膜能通过改变其动力学来适应这种变化(Xu et al., 2022)。例如, Wang等(2013)采用TIRFM和荧光相关光谱技术(fluorescence correlation spectroscopy, FCS), 在单分子水平上对拟南芥根表皮细胞质膜铵转运蛋白AMT1;3进行了活体动态分析, 发现在高铵胁迫下, AMT1;3在特定的膜微区迅速聚集, 随后通过胞吞作用进入胞质降解, 使质膜上具有转运活性的AMT1;3蛋白减少, 细胞的铵转运能力降低。Chu等(2018)对水稻(Oryza sativa)水通道蛋白OsPIPs (OsPIP1;1、OsPIP2;4和OsPIP2;5)的分布和运动状态进行了观察, 发现定位于PM的OsPIPs蛋白经过盐及渗透胁迫后, 在PM上的数量减少且运动状态发生改变, 表明在外界环境胁迫下PM蛋白可能重新定位并表现出不同的动力学特征。此外, Cui等(2018a)运用单分子成像和单颗粒追踪分析技术, 对拟南芥细胞质膜FLS2蛋白进行了时空动态分析, 发现FLS2 (FLAGELLIN-SENSITIVE 2)被特异性配体flg22激活后, 在细胞质膜上的侧向扩散显著加快, 停留时间显著缩短, 说明其运动状态与活性密切相关。综上所述, PM蛋白的分布呈现高度动态和异质性, 且环境改变或外界条件刺激可能改变原有PM蛋白的运动状态及分布模式。
膜筏(membrane raft) (大小介于10-200 nm之间的动态结构)是细胞膜上富含鞘脂类和胆固醇的一类膜微区(membrane microdomain)。膜蛋白存在于质膜上, 不同的质膜蛋白其动力学特征不同。有研究表明, 膜蛋白扩散的不均一性及表现出的受限运动均与膜筏相关(Cui et al., 2018a)。对动物细胞的研究表明, 膜筏微区能够区隔化(compartmentalize)膜蛋白的扩散, 而膜蛋白进筏和出筏的运动过程对蛋白的聚合及激活均起一定的作用(Douglass and Vale, 2005; Pinaud et al., 2009)。在植物细胞中的研究也表明, 膜筏会对质膜蛋白的运动产生影响。例如, Xue等(2018)采用双通道VA-TIRFM观察phot1-GFP和AtRem1.3-mCherry在质膜上的分布情况, 并以此来确定phot1-GFP运动是否与膜微区有关, 共定位结果显示, phot1-GFP定位在膜筏上, 当用MβCD处理后, phot1-GFP在膜上的扩散面积及速率均发生明显改变, 而添加β-谷甾醇进行回补实验时, 发现phot1动力学可以恢复, 证明膜筏确实限制phot1在膜上的运动(图1D)。同样, Hao等(2014)采用MβCD处理的方法研究膜微结构域在GFP-RbohD动力学中的作用, 发现MβCD处理可促进GFP-RbohD聚集并重新分布到团簇结构(clusters)中, 且其扩散系数发生明显改变, 表明膜微区影响GFP-RbohD的定位和动力学。此外, 为进一步研究RbohD与膜微结构域之间的关系, 他们还对RbohD与膜微结构域标记物Flot1的共定位情况进行了分析, PPI (protein proximity indexes)数据结果再次证明RbohD的横向扩散与膜微区相关。与此同时, 膜筏作为众多生物学过程的重要反应平台(reaction platform), 在胞吞以及膜信号传递过程中也发挥重要的作用。在对BRI1蛋白的运动和胞吞研究中发现, 当用外源油菜素内酯(BR)处理植物后, BRI1-GFP与AtFlot1-mCherry存在共定位现象, 这些共定位的荧光点可在质膜上共扩散, 且BR处理能增加BRI1-GFP与AtFlot1-mCherry的结合, 这些结果表明膜微区的确参与了BRI1在质膜上的胞吞过程, 并揭示了BRI1的胞吞可通过2条途径进行, 即网格蛋白介导的胞吞途径和膜筏介导的胞吞途径(Wang et al., 2015)。此外, 膜筏可通过参与FLS2的胞吞过程影响信号传递。Cui等(2018a)研究发现当固醇发生突变时, flg22诱导的FLS2胞吞途径被显著抑制, 且固醇突变体也显著影响flg22诱导的信号传递。可见, 膜微区可调控质膜蛋白在膜上的动态, 破坏膜筏将影响质膜蛋白的动力学行为, 暗示膜微区是质膜蛋白正常发挥作用所必需的重要平台。
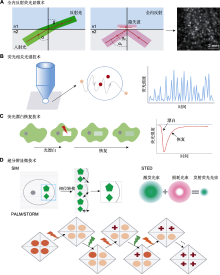
植物质膜蛋白动力学研究依赖显微观测技术。随着标记技术及显微成像系统的不断改进与发展, 显微观测的分辨率及精确度均大幅提高。全内反射荧光显微镜和荧光相关光谱法的出现不仅实现了从单层、静态、非活体到立体、动态和活体观察的飞跃, 而且使研究质膜蛋白在膜上的运动状况成为可能。新技术与各种荧光显微镜及显微技术之间的结合使用, 也将显微成像的分辨率提升至前所未有的高度。目前, 在植物质膜蛋白动力学研究中使用较为普遍的有全内反射荧光显微镜(total internal reflection fluorescence microscopy, TIRFM)、荧光相关光谱、荧光漂白恢复(fluorescence recovery after photobleaching, FRA-P)和超分辨显微等技术。下面在简要介绍这几种技术原理的基础上(图2), 对其适用对象和优缺点进行比较(表1), 同时总结其在膜蛋白动力学和植物上的应用进展, 以期更好地将显微成像技术应用于植物学领域。
(A) 由折射到全内反射(total internal reflection, TIR): 1束光在2种不同的介质中传播, 介质的折射率分别为n1和n2, 入射角为α, 由Snell定律可知, 光会发生折射进入另一种介质, 且在另一种介质中的传播角度与原介质不同, 当入射角α1等于甚至大于临界角α2时, 光就不会进入到较小折射率的介质中, 而是完全被反射到原来的介质中, 即发生全内反射; (B) 在荧光相关光谱法(FCS)中, 单个标记分子在焦体积内外的扩散会引起荧光随时间的波动, 通过将荧光涨落曲线拟合到适当的扩散模型, 可计算粒子数及其扩散系数; (C) 选择感兴趣区域(ROI), 用强激光束漂白, 并测量ROI中的荧光恢复时间; (D) 超分辨显微成像技术的原理示意图。FCS、SIM、STED、PALM和STORM同
全内反射荧光显微镜是全内反射的光学现象在光学显微镜领域的应用, 因该技术仅能照明样品表面几百纳米范围内的分子, 所以可获得高信噪比的成像效果, 及Z轴方向上的高分辨率。TIRFM经常用于检测受体与配体的互作、膜组分动力学、细胞骨架及细胞信号转导等生物学过程。但由于植物中存在细胞壁(厚约50-300 nm)的限制, 且植物细胞并不能像动物细胞那样可在培养基质或盖玻片上贴壁生长, 因此增加了一层不能确定厚度的液相基质, 入射光难以形成有效的隐失场, 从而限制了全内反射显微镜在植物细胞研究中的应用(Sundd et al., 2010; 钱虹萍等, 2021)。Wan等(2011)在全内反射荧光显微镜上安装了可调入射角度的装置, 经过理论推导和精密计算后发现, 当把入射角度调整为65.2°-72.5°时, 并将植物细胞与盖玻片紧密相贴, 入射激光可在细胞膜附近形成薄层照射, 产生约100 nm的隐失场, 从而实现在植物中对膜蛋白的动态观测, 该项技术也被称为可变角度(variable-angle)全内反射荧光显微镜术, 并应用于活体植物细胞膜蛋白的研究中(Li et al., 2011; Son et al., 2014)。例如, Yu等(2020)使用VA-TIRFM监测表皮细胞中FER-GFP的动态并分析其动力学特征, 快速碱化因子RALF1 (rapid alkalinization factor 1)处理使FER-GFP的扩散系数显著降低, 这表明RALF1处理限制了FER-GFP在PM上的扩散。Su等(2021)使用VA-TIRFM观察BSK1-GFP的定位模式, 发现BSK1-GFP荧光焦点呈点状分布, 大多数BSK1- GFP荧光点定位于PM上, 且BSK1蛋白在PM处分布不均匀, 表现出侧向和时间的动态特征。近几年, 随着显微技术的飞速发展, 研究者为获得更精确的成像效果, 常将TIRFM与其它技术结合, 如TIRF-SIM技术。TIRF-SIM法可突破传统光学显微镜衍射的极限, 获得更高的分辨率。Wollert和Langford (2022)已将TIRF-SIM应用于活细胞中肌动蛋白细胞骨架的超分辨率成像。综上所述, TIRFM技术的应用范围被逐渐拓宽, 结合其它技术可突破其衍射极限的限制, 是显微成像领域更有意义的进步。
本文的其它图/表
|
 首页
首页