|
|
||||||||
|
水稻雄性不育突变体ms102的鉴定和基因定位
植物学报
2022, 57 (1):
42-55.
DOI: 10.11983/CBB21158
水稻(Oryza sativa)隐性核雄性不育突变体是第三代杂交水稻技术的核心。为了挖掘优质雄性不育突变体, 该研究通过筛选优质籼稻黄华占(HHZ)的甲基磺酸乙酯(EMS)诱变突变体库, 获得1个雄性不育突变体ms102 (male sterility mutant 102)。该突变体营养生长正常, 但花药不开裂, 花粉败育。细胞学分析表明, 突变体花药绒毡层不能正常降解, 导致小孢子发育异常; 遗传分析表明, 该突变体的不育表型由1个已报道编码酰基转移酶的DPW2基因突变造成。研究获得了1个隐性核雄性不育突变体, 进一步证实了DPW2基因在水稻花药发育中的功能。
表3
水稻WT × ms102杂交F2代表型分离比
正文中引用本图/表的段落
利用高分辨率溶解曲线(high resolution melting, HRM)分析方法(Lochlainn et al., 2011)鉴定F2群体的基因型, 具体步骤如下: 在突变位点两侧设计PCR引物, 扩增片段约80-120 bp。PCR扩增体系: 1 μL 10× Buffer, 0.1 μL 2.5 mmol∙L-1 dNTP, 0.1 μL 10 μmol∙L-1正向引物, 0.1 μL 10 μmol∙L-1反向引物, 0.1 μL EvaGreen Dye (Li et al., 2010b), 1 μL DNA (参照CTAB法提取植株叶片DNA), 0.1 μL rTaq, 7.55 μL ddH2O, 总体积10 μL。PCR反应程序: 94°C预变性3分钟; 94°C变性30秒, 55-58°C退火30秒, 72°C延伸10秒, 40个循环; 72°C延伸1分钟; 95°C变性2分钟; 常温下复性。PCR反应完成后, 在产物里加0.1 μL低温内标(internal temperature calibrators, 纠正孔与孔之间的温度差异, 提高检测的灵敏性), 转移到Light Scanner仪器中进行扫描分型。根据分型结果, 判断F2群体该突变位点的基因型是否与表型共分离。本研究用于3个候选位点的HRM引物序列见表2。
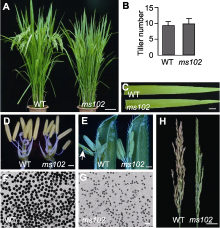
通过筛选EMS诱变的水稻突变体库, 我们获得1个完全雄性不育的突变体, 命名为ms102。该突变体植株株型、分蘖数及叶色等农艺性状均与野生型(wild type, WT)无显著差异(图1A-C)。在水稻开花期, 对突变体与野生型小花进行解剖分析, 发现与野生型相比, 突变体花药细小, 雌蕊发育正常(图1D)。花粉成熟后, 野生型的花药可以正常开裂, 但突变体的花药不能正常开裂(图1E)。对成熟期的花粉进行I2-KI染色观察, 发现突变体的花粉粒皱缩不规则, 染色表现为典败(图1F, G)。ms102突变体植株最终表现为不结实(图1H)。ms102突变体与野生型黄华占杂交的F1代所有植株育性均正常。F2代群体出现育性分离, 统计分析发现可育植株与不育植株的比例符合3:1的分离比(表3), 表明ms102突变体的雄性不育表型由单个核基因隐性突变造成。
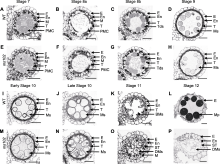
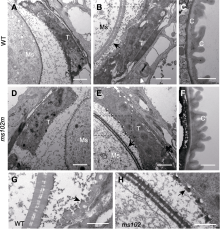
通过透射电镜观察进一步比较野生型和突变体花药的超微结构, 结果发现突变体花药发育在第9-10时期绒毡层开始出现异常, 主要表现为突变体绒毡层细胞稍有膨大(图3A, B, D, E), 乌氏体比野生型小(图3B, E, G, H), 小孢子外壁比野生型薄(图3B, E, G, H); 突变体花药外壁的角质层与野生型相比无明显差异(图3C, F)。由于花粉发育与绒毡层降解有密切关系, 绒毡层降解异常通常伴随着花粉不同程度的败育。结合突变体花药发育第1-8时期均表现正常, 我们推测, 突变基因在花药发育的第9-10时期对雄性发育起关键调控作用。
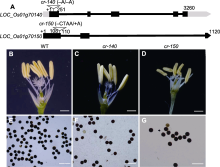
由于S3候选位点与ms102雄性不育表型紧密连锁, 我们推测控制雄性不育的基因可能在S3候选位点附近。基因组序列分析表明, S3位点下游约91 kb处存在一个已报道的雄性不育基因DPW2 (LOC_Os01g70025)。据Xu等(2017)报道, dpw2突变体的花药瘦小且为苍白色, 绒毡层降解延迟, 花粉外壁发育存在缺陷, 花粉粒形状不规则, 花粉没有活力, 同时花药外壁角质层发育受到影响。因此, 我们克隆并比较了野生型和ms102突变体中DPW2基因序列, 发现ms102突变体中DPW2基因第1 708位出现碱基替换, 导致脯氨酸突变为丝氨酸; 第1 713-1 740位共28个核苷酸缺失, 导致蛋白质翻译提前终止(图7)。此外, 我们以ms102雄性不育突变体为母本, 与dpw2杂合突变体(dpw2/+)杂交, 对F1代植株的基因型和表型进行分析, 结果发现ms102/dpw2基因型植株为雄性不育, 而ms102/+杂合体植株育性正常(图8)。以上结果表明, 控制ms102雄性不育表型的是DPW2基因上产生的新等位突变。
在前期研究中, 本课题组利用SIMM方法克隆到一系列影响水稻育性的基因(Chang et al., 2016a, 2016b, 2018, 2020; Pan et al., 2020; Peng et al., 2020; Zhang et al., 2020)。然而, 由于SIMM方法基于突变体基因组二代测序数据与参考基因组比对, 而Illumina高通量测序往往不能完全覆盖基因组序列, 造成部分信息缺失。因此获得的基因组序列无法与突变体中缺失的序列进行正确区分, 导致使用基因组重测序的方法很难识别缺失突变, 因此SIMM方法对鉴定碱基缺失突变并不敏感(Yan et al., 2017)。本研究采用SIMM方法对ms102的雄性不育突变基因进行鉴定, 发现了3个SNP候选位点, 连锁分析发现S1和S2位点都存在与突变体表型不连锁的植株, 只有S3位点与突变体表型紧密连锁。S3位于LOC_Os01g70140和LOC_Os01g70150两个基因间区, 距2个基因起始位点均在2 kb之内, 因此推测该突变可能通过影响启动子活性而影响基因功能, 对S3位点两侧基因分别敲除加以验证。然而基因敲除纯合突变体植株结实正常, 表明LOC_Os01g70140和LOC_Os01g70150不是控制ms102突变体不育表型的基因。进一步对S3位点附近的基因进行筛查, 发现距该位点约91 kb处存在1个已经报道的雄性不育基因DPW2。经基因克隆与DNA测序分析, 发现DPW2基因在ms102中发生缺失突变, 是造成ms102雄性不育的原因。因此, 利用基因组重测序、SIMM或Mut-map等方法鉴定突变位点, 在验证候选位点时, 需加大F2分离群体的数量, 对基因型和表型进行连锁分析, 并通过基因敲除和基因组片段互补等实验进行验证。此外, 特别注意的是需要对紧密连锁的候选位点附近的基因进行分析, 若发现待研究的突变体表型与已有报道的基因表型相似时, 可通过基因克隆、DNA测序对野生型和突变体的基因片段进行比较分析, 并进行遗传验证。综合以上策略, 可快速鉴定造成表型变异的突变基因位点。
本文的其它图/表
|
 首页
首页