|
|
||||||||||||||||||||||||
|
植物响应镉胁迫的生理生化机制研究进展
植物学报
2021, 56 (3):
347-362.
DOI: 10.11983/CBB20160
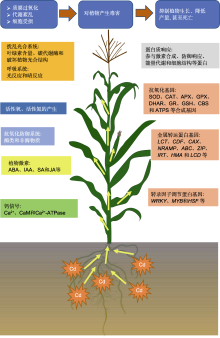
镉(Cd)是一种分布广泛且污染严重的重金属; 其毒性大, 不仅影响植物的生长发育, 而且危害人类健康。该文对植物Cd胁迫的生理生化响应方面的最新研究进展进行了总结概括。从植物光合系统、活性氧、活性氮、抗氧化防御系统、激素、钙信号、蛋白和基因等方面, 概述了植物对Cd胁迫的响应及应答机制, 探讨了植物对Cd胁迫响应机制的研究方向, 旨在为今后开展植物响应Cd胁迫的生理生化及分子机制研究提供理论依据。
表2
植物蛋白对Cd胁迫的响应
正文中引用本图/表的段落
ABA诱导的NADPH氧化酶产生的过氧化氢(H2O2)在ABA信号转导通路中发挥重要作用, 从而诱导抗氧化防御系统的启动(Hu et al., 2005)。ABA处理可抵消Cd诱导的非酶类抗氧化物质和抗氧化酶类物质的波动。例如, Li等(2014)研究表明, ABA处理显著提高了APX和POD活性, 降低了SOD和CAT活性; 使AsA水平升高, 并恢复了Cd应激引起的GSH水平降低。Bashir等(2019)研究表明, 用CdCl2溶液短期处理植物可使ABA合成基因(Glyma17G242200)表达上调, 内源性ABA浓度升高。外源ABA提高了植物螯合肽酶基因(StPCS1)的转录水平及植物螯合肽合成酶(phytochelatin synthase, PCS)活性, 且实验表明ABA参与了PCS的调控(Stroiński et al., 2010)。Fan等(2014)研究表明, 施用外源ABA可减少拟南芥中Cd的积累, 这与抑制铁调控运输因子(iron regulated transporter1, IRT1)介导的Cd吸收有关。
Cd处理的植物初级根尖和子叶中IAA浓度及分布有较大差异。Cd处理后, IAA含量显著降低(Guo et al., 2019), IAA合成基因表达下调(Bashir et al., 2019), IAA氧化酶活性增强, IAA生物合成和分解代谢基因表达改变(Hu et al., 2013)。此外, Cd胁迫下, 外源IAA能显著提高玉米幼苗根和地上部生物量, 降低地上部Cd含量, 但对根Cd含量的影响不显著, 显著降低叶片的MDA和GSH含量, 使玉米幼苗叶片SOD、POD和CAT活性显著增强(陈晶等, 2016)。当IAA合成受到抑制时, 如施加生长素信号抑制剂氯苯氧基异丁酸, 可有效抑制10 μmol·L-1 Cd诱导的根系生长和H2O2产生, 但不抑制60 μmol·L-1 Cd诱导的根系生长和H2O2产生(Tamás et al., 2012)。Cd胁迫下施加IAA, 可激活谷胱甘肽-S-转移酶(glutathione- S-transferase, GST) (在植物解毒过程中起关键作用)活性, 使植物更好地发生应激反应(Bo?ová et al., 2013)。Cd胁迫下, 施加外源IAA还可显著提高SODs基因的表达, 降低H2O2含量, 从而减少Cd对植物的毒害(Pan et al., 2020)。重金属污染条件下, 外源IAA可促进天然抗性相关巨噬细胞蛋白基因NRAMP (NATURAL RESISTANCE-ASSOCIATED MACROPHAGE PROTEIN)、IRT、重金属ATP酶家族基因HMA (heavy metal ATPase)和锌铁转运蛋白基因ZIP (zinc iron transporter protein)的表达, 提高植物的抗Cd能力(Korshunova et al., 1999; 罗莎, 2017)。
质膜上的Ca2+-ATPase能使高浓度的Ca2+进入非原生质体空间, 调节胁迫下细胞内的Ca2+平衡和维持细胞相对稳定的内环境(Snedden and Fromm, 2001; Hu et al., 2007)。赵士诚等(2008)用不同浓度的Cd处理玉米叶片, 其Ca2+-ATPase活性均表现为细胞质膜>液泡膜、内质网膜>线粒体膜。并且低浓度Cd处理下, 为保持细胞内的Ca2+平衡, 玉米叶片生物膜系统的Ca2+-ATPase活性增强; 高浓度Cd处理下, Ca2+-ATPase活性下降, 出现代谢紊乱, 玉米中下部叶变黄且生长停滞。Ca预处理可显著降低扁豆(Lablab purpureus)幼苗的Cd积累, 并通过调节H2O2酶活性和降低膜的氧化损伤减轻Cd处理的不利影响(Talukdar, 2012)。林啸等(2014)发现外源Ca处理可降低白菜(Brassica pekinensis)叶片的Cd含量, 并通过Ca2+-CaM信号途径调节酶类抗氧化剂APX、CAT和POD以及非酶类抗氧化剂GSH含量, 维持叶片ROS的平衡, 减少Cd诱导的细胞程序性死亡, 从而增强植物对Cd的耐受性。植物钙信号与抗氧化系统的关联机制值得进一步研究。
植物响应Cd胁迫的蛋白主要与抗氧化、转运和转录调控等相关(表2)。罗琼等(2015)在重金属超富集植物龙葵(Solanum nigrum)根和叶中分别鉴定出9个和6个差异表达蛋白, 涉及激素合成、防御响应、能量代谢和细胞结构等。Xu等(2017)采用定量多组分蛋白质组学方法, 对萝卜(Raphanus sativus)根系Cd胁迫下的差异表达蛋白进行了研究, 通过功能注释揭示它们主要参与碳水化合物和能量代谢、应激和防御以及信号转导过程。利用实时荧光定量PCR证实了12个基因编码其相应差异表达蛋白的表达模式, 特别是一些与碳水化合物代谢、活性氧清除、细胞转运和信号转导相关的关键蛋白参与了萝卜Cd应激反应的调控网络。Cd2+处理萝卜根后激活了几个关键信号分子(Ca2+、CaM、IAA和JA)和金属转运蛋白基因(ZIP、HMA、ABC和IRT)的表达, 减轻Cd胁迫并重建萝卜体内氧化还原稳态(Xu et al., 2017; 2020)。
根系从土壤中吸收Cd及其在植物体内运输需要多个不同的重金属转运体家族(如低亲和性阳离子转运蛋白(low-affinity cation transporter, LCT)、阳离子扩散促进剂(cation diffusion facilitator, CDF)和离子交换剂(cation exchanger, CAX))参与(孙瑞莲和周启星, 2005; Yuan et al., 2012; 袁连玉等, 2017)。这些转运体介导了植物中过渡金属离子(如Zn2+、Cd2+、Co2+、Ni2+和Mn2+)从细胞质到外腔室的空泡隔离、储存和运输过程, 被命名为重金属忍耐蛋白(metal tolerance protein, MTP) (Blaudez et al., 2003; Korenkov et al., 2009)。NRAMP家族在生物体中Mn2+、Fe2+、Zn2+和Cd2+的细胞膜转运中发挥重要作用。NRAMP家族成员将二价金属阳离子运输到细胞质中, NRAMP3在鼠耳芥(A. halleri)和天蓝遏蓝菜(Thlaspi caerulescens)根中高表达, NRAMP1、NRAMP3和NRAMP5在天蓝遏蓝菜茎和叶中高表达(Kr?mer et al., 2007)。Zhang等(2020a)在Cd超积累植物东南景天(Sedum alfredii)中发现, SaNRAMP1过表达显著增加了烟草(Nicotiana tabacum)茎和叶中的Cd浓度。Zhang等(2020b)在平邑甜茶(Malus hupehensis)中发现, MhNRAMP1的表达蛋白是一种位于细胞膜的疏水性跨膜蛋白, Cd胁迫和Ca2+缺乏诱导MhNRAMP1在根中表达。MhNRAMP1过表达可增加酵母中Cd浓度, 增强其对Cd的敏感性。Cd胁迫下, MhNRAMP1过表达的转基因烟草和苹果(M. Pumila)愈伤组织生长状况比野生型(WT)差。MhNRAMP1通过介导Cd的吸收和积累及刺激液泡加工酶, 加速Cd诱导的细胞死亡。PCS和尼克烟酰胺是植物体内常见的重金属螯合剂, YSL (yellow stripe1-like)基因家族在螯合金属的转运过程中发挥重要作用(Curie et al., 2009)。Feng等(2017)发现SnYSL3编码一种等离子体局部转运蛋白, 转运多种金属-烟酰胺复合物。Cd胁迫可上调SnYSL3的表达, 表明SnYSL3在Cd胁迫中起重要作用。
重金属ATP酶基因家族(HMA)已被证明在不同的植物种类中行使特定的功能, 不同的HMA在不同组织中表达, 最终导致功能的多样性。HvHMA1参与Zn以及Cd向大麦(Hordeum vulgare)籽粒的转运(Mikkelsen et al., 2012)。HMA2和HMA4可将Cd装载到木质部, 以便从根运输至茎和叶, OsHMA2与水稻维管组织中的锌装载及液泡膜定位有关(Yamaji et al., 2013)。液泡膜上的OsHMA3将Cd转移至根系液泡, OsHMA4则将Cu转运至根系液泡, 限制Cd从根向茎以及叶的转移, 从而减少Cd在种子中的积累(Miyadate et al., 2011; Huang et al., 2016)。TcHMA3在叶片的液泡中表达, 将Cd隔离在根和叶的液泡中, 最终导致Cd的超积累, 增强对Cd的耐受性(Ueno et al., 2011)。AtHMA3在拟南芥中过表达导致Cd的积累量比野生型高2-3倍(Morel et al., 2009)。这些转运蛋白为植物的基因工程提供了有用的工具, 增强了植物对金属的耐受性和积累能力, 继续推进它们的协同作用研究具有重要意义。
MT: 金属硫蛋白; PC: 螯合肽; ROS: 活性氧; RNS: 活性氮.GSH、SOD、CAT、APX、GPX、DHAR、GR、CBS和ATPS同 Melatonin confers cadmium tolerance by modulating critical heavy metal chelators and transporters in radish plants 2 2020 ... 植物响应Cd胁迫的蛋白主要与抗氧化、转运和转录调控等相关(
本文的其它图/表
|
||||||||||||||||||||||||
 首页
首页