|
|
||||||||||||||||||||||||||||||
|
植物响应镉胁迫的生理生化机制研究进展
植物学报
2021, 56 (3):
347-362.
DOI: 10.11983/CBB20160
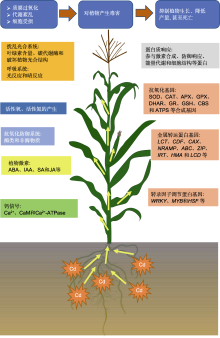
镉(Cd)是一种分布广泛且污染严重的重金属; 其毒性大, 不仅影响植物的生长发育, 而且危害人类健康。该文对植物Cd胁迫的生理生化响应方面的最新研究进展进行了总结概括。从植物光合系统、活性氧、活性氮、抗氧化防御系统、激素、钙信号、蛋白和基因等方面, 概述了植物对Cd胁迫的响应及应答机制, 探讨了植物对Cd胁迫响应机制的研究方向, 旨在为今后开展植物响应Cd胁迫的生理生化及分子机制研究提供理论依据。
表1
植物激素对Cd胁迫的响应
正文中引用本图/表的段落
Cd胁迫下, 抗逆相关的植物激素(如ABA、IAA、SA和JA)均会发生显著变化, 且影响抗氧化物质合成相关基因和运输蛋白基因的表达(表1)。多数情况下Cd胁迫可引起植物体内IAA、SA、ABA和JA含量变化(Liu et al., 2016; Bashir et al., 2019; Song et al., 2019)。添加适量外源ABA、IAA、SA和JA等可缓解Cd胁迫对植物的危害。
ABA诱导的NADPH氧化酶产生的过氧化氢(H2O2)在ABA信号转导通路中发挥重要作用, 从而诱导抗氧化防御系统的启动(Hu et al., 2005)。ABA处理可抵消Cd诱导的非酶类抗氧化物质和抗氧化酶类物质的波动。例如, Li等(2014)研究表明, ABA处理显著提高了APX和POD活性, 降低了SOD和CAT活性; 使AsA水平升高, 并恢复了Cd应激引起的GSH水平降低。Bashir等(2019)研究表明, 用CdCl2溶液短期处理植物可使ABA合成基因(Glyma17G242200)表达上调, 内源性ABA浓度升高。外源ABA提高了植物螯合肽酶基因(StPCS1)的转录水平及植物螯合肽合成酶(phytochelatin synthase, PCS)活性, 且实验表明ABA参与了PCS的调控(Stroiński et al., 2010)。Fan等(2014)研究表明, 施用外源ABA可减少拟南芥中Cd的积累, 这与抑制铁调控运输因子(iron regulated transporter1, IRT1)介导的Cd吸收有关。
Cd胁迫通常促进JA合成基因(Bashir et al., 2019)和抗氧化酶合成基因的转录(Keunen et al., 2013)。低浓度JA可缓解Cd胁迫。例如, 用20 μmol·L-1 JA处理可有效改善Cd诱导的氧化应激, 使硫代巴比妥酸反应物质、H2O2和O2-.的积累减少, 使GSH、CAT、SOD及血红素氧合酶(HO)活性增强(Noriega et al., 2012; Adhikari et al., 2018)。研究发现, 番茄(Solanum lycopersicum) JA缺失突变体对Cd的敏感性增强, 与光合作用相关参数(包括总叶绿素、光系统II实际效率、可变叶绿素荧光与最大荧光比及净光合速率)值降低; 相对水含量、可溶性糖和脯氨酸含量及淀粉的积累减少; 氧化应激和抗氧化防御作用(包括H2O2与MDA生成、电解质渗漏、SOD、APX和CAT活性)减弱(Zhao et al., 2016)。外源茉莉酸甲酯(MeJA)通过降低拟南芥根系和茎叶中的Cd浓度, 下调参与Cd吸收和运输的AtIRT1、AtHMA2和AtHMA4基因表达, 缓解Cd引起的新叶毒害。JA合成关键基因AtAOS突变上调AtIRT1、AtHMA2和AtHMA4等转运蛋白基因表达, 增加根和芽中的Cd浓度, 提高拟南芥对Cd的敏感性。此外, MeJA降低Cd胁迫的拟南芥根尖中的NO水平(Lei et al., 2020)。镉胁迫下植物激素ABA、IAA、SA和JA之间的相互作用值得深入探索。
根系从土壤中吸收Cd及其在植物体内运输需要多个不同的重金属转运体家族(如低亲和性阳离子转运蛋白(low-affinity cation transporter, LCT)、阳离子扩散促进剂(cation diffusion facilitator, CDF)和离子交换剂(cation exchanger, CAX))参与(孙瑞莲和周启星, 2005; Yuan et al., 2012; 袁连玉等, 2017)。这些转运体介导了植物中过渡金属离子(如Zn2+、Cd2+、Co2+、Ni2+和Mn2+)从细胞质到外腔室的空泡隔离、储存和运输过程, 被命名为重金属忍耐蛋白(metal tolerance protein, MTP) (Blaudez et al., 2003; Korenkov et al., 2009)。NRAMP家族在生物体中Mn2+、Fe2+、Zn2+和Cd2+的细胞膜转运中发挥重要作用。NRAMP家族成员将二价金属阳离子运输到细胞质中, NRAMP3在鼠耳芥(A. halleri)和天蓝遏蓝菜(Thlaspi caerulescens)根中高表达, NRAMP1、NRAMP3和NRAMP5在天蓝遏蓝菜茎和叶中高表达(Kr?mer et al., 2007)。Zhang等(2020a)在Cd超积累植物东南景天(Sedum alfredii)中发现, SaNRAMP1过表达显著增加了烟草(Nicotiana tabacum)茎和叶中的Cd浓度。Zhang等(2020b)在平邑甜茶(Malus hupehensis)中发现, MhNRAMP1的表达蛋白是一种位于细胞膜的疏水性跨膜蛋白, Cd胁迫和Ca2+缺乏诱导MhNRAMP1在根中表达。MhNRAMP1过表达可增加酵母中Cd浓度, 增强其对Cd的敏感性。Cd胁迫下, MhNRAMP1过表达的转基因烟草和苹果(M. Pumila)愈伤组织生长状况比野生型(WT)差。MhNRAMP1通过介导Cd的吸收和积累及刺激液泡加工酶, 加速Cd诱导的细胞死亡。PCS和尼克烟酰胺是植物体内常见的重金属螯合剂, YSL (yellow stripe1-like)基因家族在螯合金属的转运过程中发挥重要作用(Curie et al., 2009)。Feng等(2017)发现SnYSL3编码一种等离子体局部转运蛋白, 转运多种金属-烟酰胺复合物。Cd胁迫可上调SnYSL3的表达, 表明SnYSL3在Cd胁迫中起重要作用。
重金属ATP酶基因家族(HMA)已被证明在不同的植物种类中行使特定的功能, 不同的HMA在不同组织中表达, 最终导致功能的多样性。HvHMA1参与Zn以及Cd向大麦(Hordeum vulgare)籽粒的转运(Mikkelsen et al., 2012)。HMA2和HMA4可将Cd装载到木质部, 以便从根运输至茎和叶, OsHMA2与水稻维管组织中的锌装载及液泡膜定位有关(Yamaji et al., 2013)。液泡膜上的OsHMA3将Cd转移至根系液泡, OsHMA4则将Cu转运至根系液泡, 限制Cd从根向茎以及叶的转移, 从而减少Cd在种子中的积累(Miyadate et al., 2011; Huang et al., 2016)。TcHMA3在叶片的液泡中表达, 将Cd隔离在根和叶的液泡中, 最终导致Cd的超积累, 增强对Cd的耐受性(Ueno et al., 2011)。AtHMA3在拟南芥中过表达导致Cd的积累量比野生型高2-3倍(Morel et al., 2009)。这些转运蛋白为植物的基因工程提供了有用的工具, 增强了植物对金属的耐受性和积累能力, 继续推进它们的协同作用研究具有重要意义。
MYB转录因子控制发育和代谢, 是生物和非生物胁迫反应调控网络中的关键因素(Dubos et al., 2010)。Cd诱导拟南芥氧化应激产生ROS, 而MYB4可激活抗氧化防御系统, 提高SOD和POD等抗氧化酶活性, 并增强PCS和MTs基因的表达, 从而减轻ROS对细胞的损害(Agarwal et al., 2020)。Zhang等(2019a)研究发现, MYB49通过直接与基本螺旋-环-螺旋(helix loop helix, HLH)的转录因子bHLH38和bHLH101启动子结合, 正向调控其表达, 导致IRT1被激活, 而IRT1编码一种参与Cd摄取的金属转运蛋白。MYB49还与重金属相关的异丙烯化植物蛋白(heavy metal-associated isoprenylated plant proteins, HIPP) HIPP22和HIPP44的启动子区域结合, 导致它们表达上调, Cd的积累增加。此外, 作为控制Cd在植物细胞中吸收和积累的反馈机制, Cd诱导ABA上调ABI5的表达, 其蛋白产物与MYB49互作, 并阻止其与下游基因启动子结合, 从而减少Cd的积累。ABA信号控制Cd在植物中吸收和积累的分子反馈机制与MYB49蛋白调控途径具有一定的关系。
AsA: 抗坏血酸; POD: 过氧化物酶; GST: 谷胱甘肽-S-转移酶; NR: 硝酸还原酶; GS: 谷氨酰胺合成酶; GOGAT: 谷氨酸合成酶.ABA、IAA、SA、JA、GSH、SOD、CAT和APX同 Effects of calcium application on activities of membrane transporters in Panax notoginseng under cadmium stress 1 2021 ... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害( RETRACTED: the interplay between reactive oxygen and nitrogen species contributes in the regulatory mechanism of the nitro-oxidative stress induced by cadmium in Arabidopsis 1 2018 ... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害( Improved short-term drought response of transgenic rice over-expressing maize C4 phosphoenolpyruvate carboxylase via calcium signal cascade 1 2017 ... Ca2+作为细胞功能的第二信使, 参与调节植物各种逆境下的重要生理生化过程.钙信使系统包括Ca2+、钙调蛋白(CaM)和钙泵(Ca2+-ATPase).CaM是Ca2+的感受器, Ca2+-ATPase决定着Ca2+的跨膜运输( Role of salicylic acid in resistance to cadmium stress in plants 4 2016 ... Cd胁迫下, 抗逆相关的植物激素(如ABA、IAA、SA和JA)均会发生显著变化, 且影响抗氧化物质合成相关基因和运输蛋白基因的表达(
本文的其它图/表
|
 首页
首页