|
|
|||||||||||||||||||||||||||||||||||||||||||
|
辣椒R2R3-MYB转录因子家族的全基因组鉴定与比较进化分析
植物学报
2021, 56 (3):
315-329.
DOI: 10.11983/CBB20143
MYB转录因子作为植物中最大的转录因子家族之一, 参与植物的生长、代谢、抵御生物和非生物胁迫等多种生理生化过程。R2R3-MYB是MYB转录因子家族的主要存在形式。辣椒是具有重要经济价值的蔬菜作物, 其R2R3-MYB转录因子缺乏系统的研究。从一年生辣椒(Capsicum annuum)、浆果状辣椒(C. baccatum)和中国辣椒(C. chinense)基因组中分别鉴定出94、92和94个R2R3-MYB基因, 基于系统发育关系将其分为28个亚族。共线性分析表明, 3种辣椒间存在73组直系同源R2R3-MYB基因, 一年生辣椒、浆果状辣椒和中国辣椒分别存在5、4和2个特有的R2R3-MYB基因。鉴定出12对重复基因, 其中8对是串联重复基因, 它们在3种辣椒分化前就已经存在。比较基因组学分析表明, 在辣椒进化过程中同源R2R3-MYB转录因子发生了功能分化。组织表达分析表明, 辣椒R2R3-MYB基因主要有3种表达特征: 在根、叶、茎和花中均高表达, 如CaMYB13/CbMYB12/CcMYB13; 仅在花中高表达, 如CaMYB93/CbMYB86/CcMYB12; 仅在根中高表达, 如CaMYB48/CbMYB47/CcMYB51。研究结果为深入揭示R2R3-MYB转录因子在辣椒生长发育中的生物学功能奠定了基础。
表1
本研究使用的引物序列
正文中引用本图/表的段落
采用实时荧光定量PCR (qRT-PCR)检测辣椒MYB基因的组织表达, 根据一年生辣椒MYB基因CDS序列设计qRT-PCR引物(表1)。实验材料是由安徽科技学院辣椒课题组提供的一年生辣椒自交系(编号为G7②), 于2019年12月上旬播种于安徽科技学院种植科技园。采集生长6周植株的根、茎和叶组织和盛花期开放的花组织, 利用Trizol试剂(Life Technologies, 美国)提取各组织的RNA, 使用HiProTM (H-) 1st Strand cDNA Synthesis Kit with gDNA Eraser (普鲁顿, 北京)反转录获得cDNA, 并用SuperReal PreMix Plus (SYBR Green) (天根, 北京)试剂盒进行定量, PCR仪为ABI ViiA7实时荧光定量PCR仪(Life Technologies, 美国)。以CaUBI-3基因作为内参基因。反应体系总体积为20 μL, 包括1 μL cDNA, 上、下游引物各0.5 μL, 10 μL SYBR, 用ddH2O补足体积。反应程序为: 95°C预变性30秒; 95°C变性10秒, 60°C退火30秒, 72°C延伸30秒, 40个循环。实验重复3次。反应结束后分析荧光值变化曲线及熔解曲线。基因相对表达量采用2-ΔΔCT法计算(Livak and Schmittgen, 2001)。
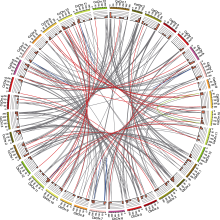
通过BLASTP比对筛选, 从一年生辣椒、浆果状辣椒和中国辣椒中分别鉴定出94、92和94个MYB转录因子基因, 它们都只包含R2和R3 MYB结构域(图1)。根据R2R3-MYB基因在染色体上的分布, 按从大到小的顺序对基因进行命名, 并利用氨基酸序列对所有R2R3-MYB转录因子进行理化性质分析(附表1)。结果表明, CaMYB、CbMYB和CcMYB蛋白大小分别介于118-539、173-545和174-563个氨基酸之间; 分子量大小分别介于13.96-60.08、20.02-60.03和20.07- 62.62 kDa之间; 等电点在4.61-10.3、4.33-10.28和4.53-10.65之间。蛋白定位预测结果显示, R2R3-MYB蛋白主要定位在细胞核, 少数蛋白定位在细胞外。
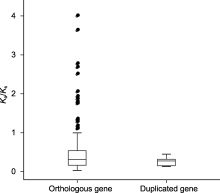
通过计算非同义替换率(Ka)和同义替换率(Ks)可以评估3种辣椒中同源基因的进化模式。Ka/Ks>1表示受到正选择, Ka/Ks<1表示受到纯化选择, Ka/Ks=1表示基因中性进化(Yadav et al., 2015)。本研究表明, 所有直系同源基因中有17对直系同源基因(CaMYB7/ CcMYB7、CaMYB9/CbMYB8、CaMYB11/CbMYB55、CaMYB18/CbMYB91、CaMYB29/CcMYB27、CaMYB30/CbMYB30、CaMYB37/CbMYB37、CaMYB37/ CcMYB38、CaMYB58/CbMYB57、CaMYB63/CcMYB63、CaMYB67/CcMYB66、CaMYB90/CcMYB81、CaMYB94/CbMYB87、CbMYB8/CcMYB10、CbMYB30/CcMYB29、CbMYB43/CcMYB50和CbMYB74/Cc MYB81)的Ka/Ks>1 (图6), 说明这些基因对受到正选择, 暗示这17对直系同源基因可能存在功能上的分化。有1对同源基因(CbMYB4/CcMYB4) Ka/Ks=1, 可能是中性进化。其余同源基因对和12对重复基因的Ka/Ks均小于1, 说明这些基因很可能受到纯化选择。
黑色点表示非同义替换率(Ka)/同义替换率(Ks)>1的基因对。
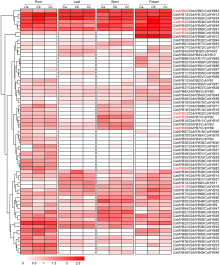
基于73组直系同源基因的转录组数据, 我们发现不同辣椒的R2R3-MYB基因在4种组织中的表达具有明显差异(图7)。综合分析发现主要有3种表达模式: (1) 在根、叶、茎和花4种组织中均高表达, 但表达量不尽相同, 如CaMYB13/CbMYB12/CcMYB13、CaMYB34/CbMYB92/CcMYB34和CaMYB35/CbMYB29/CcMYB35, 表明这些基因可能在调控辣椒生长发育过程中起重要作用; (2) 在花中特异或高表达的基因, 如CaMYB93/CbMYB86/CcMYB12; (3) 在根中特异或高表达的基因, 如CaMYB48/CbMYB47/ CcMYB51和CaMYB80/CbMYB78/CcMYB82, 这些基因可能参与相应组织发育的某些生物学过程。此外, 大多数直系同源基因在3种辣椒中的表达特征类似, 但还存在一些直系同源基因在同一组织中表达模式不同。例如, CaMYB31在所有组织中均不表达, 而它的直系同源基因CbMYB27和CcMYB31在根中表达, 表明这些直系同源基因可能发生了功能分化。
R2R3-MYB基因在辣椒12条染色体上的数目不尽相同, 但它们主要位于每条染色体的长臂或短臂末端(附表1; 图5)。3种辣椒直系同源基因在染色体上的位置很接近, 锚定在高度保守的共线性区块上。研究发现, 浆果状辣椒中3号染色体长臂与5号染色体短臂发生了易位, 3号染色体短臂和9号染色体长臂发生了易位(Kim et al., 2017), 位于这些位点的R2R3- MYB基因也发生了变化。例如, CbMYB24位于浆果状辣椒3号染色体上, 它的同源基因CcMYB36位于中国辣椒5号染色体上。此外, 还有少量直系同源R2R3- MYB基因位于不同的染色体上。例如, CbMYB38/ CcMYB42位于6号染色体, 其直系同源基因CaMYB4位于1号染色体; CaMYB10/CcMYB9位于1号染色体, 其直系同源基因CbMYB56位于8号染色体, 这可能是由染色体结构变异所致。基于已发布的基因组信息, 3种辣椒中有21个R2R3-MYB基因尚未定位到染色体上, 但和它们对应的直系同源基因却已定位到染色体上。导致这一现象的原因不排除基因组组装错误, 后续可通过进一步精细测序以及构建高质量辣椒基因组图谱来消除。直系同源基因和串联重复基因的Ka/Ks分析结果(图6)表明, 有17对直系同源基因可能受到正选择, 暗示着它们的功能可能发生了变化。其中, CaMYB37/CbMYB37、CaMYB37/CcMYB38的Ka/Ks均大于1, 但CbMYB37/CcMYB38的Ka/Ks却小于1, 表明CaMYB37和它的直系同源基因CbMYB37/CcMYB38之间可能出现了功能分化。CbMYB8和CbMYB30也有类似的进化模式。
附表1 一年生辣椒R2R3-MYB蛋白基本信息
本文的其它图/表
|
 首页
首页