木本植物性别决定基因研究进展
陆静, 陈赢男, 尹佟明
植物学报
2021, 56 ( 1):
90-103.
DOI: 10.11983/CBB20123
雌雄异株植物是研究性别决定遗传机制及性染色体起源与进化的理想材料, 而克隆性别决定基因是解析性别决定遗传机制的关键。木本植物中有丰富的雌雄异株植物, 且包括2种相反的性别决定系统: XY型(雌株为同配型的XX, 雄株为异配型的XY)和ZW型(雌株为异配型的ZW, 雄株为同配型的ZZ)。此外, 不同性别植株的经济价值也有所不同。在木本植物中开展性别决定机制研究不仅具有重要的理论意义, 还具有较高的生产应用价值。随着大规模基因测序技术的快速发展, 越来越多的木本植物性别决定基因被鉴定和克隆, 为解析雌雄异株植物性别决定机制和性染色体的演化过程提供了有力的实验证据。该文详细总结了近年来木本植物性别决定基因研究的重要进展, 并展望了未来的研究方向及发展趋势。
| 类型 | 种名 | 性别决定系统 | 性别决定
基因 | 基因注释 | 在性别分化中的作用 | 参考文献 | | 隐性雌雄异株(I型单性花) | 君迁子(Diospyros lotus) | XY | OGI | Y染色体特异的21 bp小RNA | 雄株中抑制雌蕊发育 | Akagi et al., 2014 | | 猕猴桃(Actinidia spp.) | XY | SyGI | Type-C类型细胞分裂素响应调节因子 | 雄株中抑制雌蕊发育 | Akagi et al., 2018 | | FrBy | 类成束阿拉伯半乳糖蛋白(fasciclin-like arabinogalactan proteins, FLAs)是一类广泛分布于植物体内的富含羟脯氨酸的糖蛋白 | 雄株中激活雄蕊发育 | Akagi et al., 2019 | | 葡萄(Vitis vinifera) | XY | APT3/APRT3 | 腺嘌呤磷酸核糖转移酶 | 推测在雄株抑制心皮发育 | Fechter et al., 2012; Coito et al., 2017 | | INP1 | 功能未知蛋白 | 推测与花粉萌发孔发育相关 | Massonnet et al., 2020 | | 番木瓜(Carica papaya) | XY | CpSVP | MADS-box转录因子 | 功能未知 | Urasaki et al., 2012; Lee et al., 2018 | | CpSERK | 体细胞胚发生相关类
受体蛋白激酶 | | CpCAF1AL | 组蛋白伴侣, 染色质组装因子 | 椰枣
(Phoenix dactylifera) | XY | CYP703 | 细胞色素蛋白 | 推测促进雄蕊发育 | Torres et al., 2018 | | GPAT3 | 甘油-3-磷酸酰基转移酶 | | LOG-like | 细胞分裂素激活酶 | 推测抑制雌蕊发育 | | 完全雌雄异株(II型单性花) | 美洲黑杨
(Populus deltoides) | XY | FERR-R | Y染色体特异小RNA | 抑制雌蕊发育 | Xue et al., 2020 | | MSL | Y染色体特异lncRNA | 促进雄蕊发育 | | 毛果杨(P. trichocarpa) | XY | ARR17 | Type-A类型细胞分裂素响应调节因子 | 促进雌蕊发育 | Müller et al., 2020 | | 欧洲山杨(P. tremula) | XY | ARR17 | | 胡杨(P. euphratica) | XY | RR | Type-A类型细胞分裂素响应调节因子 | Yang et al., 2020 |
View table in article
表1
目前已鉴定的木本植物性别决定基因
正文中引用本图/表的段落
柿属(Diospyros)植物不仅染色体倍性复杂, 性别类型也较为丰富, 存在雌株、雄株和雌雄异花同株等多种类型(Yonemori et al., 1993)。近年来, 柿雌雄花性别分化的调控机理研究取得了重要进展。柿属二倍体植物君迁子(D. lotus)为雌雄异株, 成熟雄花3朵合生、呈聚伞状花序, 雌花1朵单生, 其性别决定系统为XY型(Akagi et al., 2013)。在没有参考基因组序列的情况下, 研究人员对君迁子种内杂交F1代个体(32雌株和25雄株)分别进行基因组从头测序, 根据雌、雄性别将测序数据合并建成2个序列库, 通过对这2个序列库进行K-mer比较分析, 筛选出雄株特异的K-mer片段, 进行局部组装, 最终得到Y染色体上约1 Mb的雄株特异区间。结合雌雄花芽转录组差异表达分析, 结果表明该区间含有22个性别决定候选基因, 其中仅有1个基因(OGI)只在雄花中表达。此外, 通过差异表达分析还检测到1个位于常染色体上、仅在雌花中表达的基因(MeGI)。序列分析显示, OGI基因能够转录形成发卡结构, 产生小RNA分子; MeGI基因编码HD-Zip蛋白(同源异型-亮氨酸拉链蛋白), 属于高等植物特有的转录因子, 且OGI产生的小RNA能够介导MeGI基因沉默, 这也是雄花中未检测到MeGI基因表达的原因。过表达MeGI基因的拟南芥(Arabidopsis thaliana)和烟草(Nicotiana tabacum)出现雄蕊缺损、变短和花粉萌发率降低等表型, 表明MeGI基因能够抑制雄蕊发育。由此证明, 位于Y染色体特异片段上的OGI基因决定君迁子的性别, 该基因通过产生小 RNA抑制MeGI基因在雄株中表达, 从而促进雄蕊发育; 而雌株由于没有OGI基因, MeGI基因能够正常表达, 导致雌花中的雄蕊退化(Akagi et al., 2014)。随着君迁子雄株全基因组序列图谱的完成, OGI和MeGI基因的起源以及柿属植物性别决定系统的进化机制得到进一步明晰。Ks分析表明, 君迁子基因组在进化过程中经历了1次柿属植物特有的全基因组重复事件(Dd-α WGD), MeGI (Chr. 13)由其旁系同源基因SiMeGI (Chr. 4)经过全基因组重复或片段重复而来, MeGI和SiMeGI分化后演化出新的功能, 能够抑制雄蕊发育, 而SiMeGI仍保留祖先基因的功能, 调控花器官发育但不影响性别分化(Akagi et al., 2020)。基于以上发现, Akagi等(2020)提出柿属植物从雌雄同株向雌雄异株的演化模型: 在柿子祖先基因组中仅有SiMeGI基因, 其经历Dd-α WGD产生MeGI, MeGI在演化过程中获得新的功能, 导致雌株出现; MeGI经过片段重复产生OGI, OGI反过来抑制MeGI的表达, 导致雄株产生。因此, 位于Y染色体特异片段上的OGI通过调控MeGI基因表达与否决定君迁子雌雄性别。MeGI是1个重要的整合器, 它通过整合影响下游靶基因的表达进而影响雄蕊或雌蕊的发育。综合利用雌雄花芽转录组差异表达和共表达网络, Yang等(2019)预测了MeGI基因下游调控网络。该基因通过KNOX/ OFP/GRF激活细胞分裂素/生长素/赤霉素途径相关基因促进雌蕊发育; 同时通过SVP/SOC抑制PI的表达来抑制雄蕊发育。
MeGI基因对性别分化的调控作用在不同柿属植物中的保守性较高。六倍体栽培柿(D. kaki)一般仅开雌花, 性染色体组成为XXXXXX; 或者雌雄异花同株, 性染色体组成为XXXXXY/XXXXYY (简称Dka-□)。仅开雌花的栽培柿中不具有OGI基因, 能够检测到MeGI基因, 其性别决定机制与君迁子雌株相同。雌雄异花同株的栽培柿同时含有OGI (Y染色体特异)和MeGI基因, 但OGI基因在雌花和雄花中均不表达; 而MeGI仅在雌花中表达, 在雄花中不表达。研究发现, 在Dka-□的OGI启动子区域有1个268 bp的类SINE (short interspersed nuclear element-like)反转录转座子插入, 且SINE-like元件在雌、雄花和营养器官均呈现高度甲基化, 这些因素导致OGI沉默; MeGI基因启动子和编码序列在Dka-□的雄花中高度甲基化, 而在Dka-□的雌花中甲基化水平较低, 这是导致MeGI仅在雌花中表达的原因。与君迁子相比, D. kaki通过更灵活的甲基化修饰影响花的性别, 且这种表观调控方式可以发生反转(Akagi et al., 2016)。在果树生产中对雌株需求明显大于雄株, 性别决定基因的克隆为柿树的早期性别鉴定提供了有利的分子工具。例如, Zhang等(2016)利用OGI基因开发了雄株特异的分子标记, 该标记不仅能准确鉴别D. kaki杂交子代性别, 在另外9种柿属植物中也具有很好的通用性。
在已定位的葡萄性别决定区间(sex determination region, SDR)基础上, Massonnet等(2020)利用单分子实时(single-molecule real-time, SMRT)测序技术和FALCON-Unzip组装工具, 重建了11种不同性别葡萄的SDR单倍型序列, 并通过比较不同单倍型SDR的结构变异, 分析群体中与性别关联的多态性位点, 同时结合3种花型花芽的转录组数据, 提出INP1是葡萄性别决定的候选基因。该基因在雌株中显著高表达, 但雄株和雌雄同株葡萄中具备有功能的INP1完整序列, 雌株INP1基因存在8 bp缺失, 导致移码突变和编码提前终止, 推测该基因功能缺失是导致雌花花粉畸形的原因。INP1同源基因在拟南芥花粉萌发孔的形成中发挥重要作用(Dobritsa and Coerper, 2012)。此外, Massonnet等(2020)还提出葡萄从雌雄同花祖先进化为雌雄异株植物, 以及由雄株性别决定位点间的稀有重组事件导致雌雄同花葡萄再次产生的进化模型。目前, 关于葡萄性别决定基因的研究提出多个候选基因, 但大部分处于生物信息学预测阶段, 尚缺乏对基因功能进行验证的实验证据。
番木瓜性别决定机制曾存在多种假说(Ming et al., 2007), 目前被普遍接受的是Horovitz和Jiménez (1967)根据属间杂交结果提出的XX-XY模型, 认为番木瓜雌株、雄株和两性株的基因型分别是XX、XY和XY2, Y2染色体由Y突变而来, Y和Y2染色体上均含有致死因子, 因此不存在YY、YY2和Y2Y2基因型的后代。由于雄株与两性株的Y染色体相似性很高(Liu et al., 2004), Ming等(2007)用Yh替代Y2, 用XYh表示两性株基因型。染色体序列比对和共线性分析显示, Y和Yh相似性高达99.60%。进一步分析发现Yh起源于Y染色体, 在约4 000年前完成分化, 在番木瓜长期的人工驯化过程中Yh被保留和固定下来(VanBuren et al., 2015)。精细定位和FISH结果显示, 番木瓜性别决定基因位于LG1着丝粒附近约8 Mb的区间内, 该区间重组抑制严重, 且富含重复序列和转座子, 基因克隆难度较高(Yu et al., 2007; Wang et al., 2012)。近年来, 基于转录组测序多个研究团队相继报道了番木瓜性别决定候选基因。Urasaki等(2012)通过转录组差异表达分析, 发现与序列标签Cp2671对应的基因编码1个MADS-box转录因子, 与拟南芥调控花期的转录因子SVP存在85%的序列相似性。SVP-like基因是Y和Yh染色体特异基因, 仅在雄花和两性株中表达, 由此提出SVP-like基因是番木瓜性别决定候选基因。通过比较MSY (male-specific region of Y)和HSY (hermaphrodite-specific region of Yh)的序列, 发现Y染色体上的SVP-like基因编码1个完整蛋白, 但Yh上SVP-like基因内有转座子插入, 编码不完整蛋白(Ueno et al., 2015)。基于基因表达差异和选择性剪切(alternative splicing, AS)分析, Lee等(2018)同样认为CpSVPL (SVP-like)是番木瓜性别决定基因, 他们还提出另外2个性别相关基因CpSERK和CpCAF1AL。其中, CpSERK基因在不同性别花中表达量差异不显著, 但存在AS; CpCAF1AL在雄花和正常发育的两性花中表达量较高, 不存在AS, 在发育不正常的两性花中表达量较低且存在AS。Zerpa-Catanho等(2019)通过比较3种不同性别和不同发育时期番木瓜花RNA-seq数据, 发现CpMS1仅在花器官中表达, 且在雄花和两性花发育前期表达量较高, 该基因在拟南芥、大麦(Hordeum vulgare)和水稻(Oryza sativa)等植物中的同源基因功能相似, 能够调控花粉发育, 但由于CpMS1基因位于2号染色体, 且3种性别的番木瓜均含有该基因, 因此该基因不具有性别决定功能。最近, Zhou等(2019)通过对不同季节雌、雄花全基因组甲基化水平和转录组差异表达基因进行分析, 发现雄花基因组甲基化水平低于雌花, 常染色体上CpSVP和CpAP1基因的去甲基化可能是导致雄花比雌花早开放的原因。此外, 他们还发现CpARR5基因在雄花中的表达量和甲基化水平均较雌花低, 在检测到的10个与激素信号转导相关的差异表达基因中, CpARR5基因是唯一存在甲基化修饰位点差异的基因, 推测甲基化介导的植物激素稳态的改变可能影响番木瓜性别。
综上, 大部分杨树性别决定系统为XY型, 而大部分柳树性别决定系统为ZW型。比较基因组学显示, 杨树和柳树起源于共同的祖先, 基因组具有高度的共线性关系(Tuskan et al., 2006; Dai et al., 2014)。但它们的性染色体分别由不同的常染色体进化而来: 杨树的性染色体是19号染色体, 而柳树的性染色体是15号染色体(Hou et al., 2015)。细胞学研究表明, 杨树和柳树的性染色体均未出现形态上的差异, 尚处于进化早期阶段(Peto, 1938)。由此可见, 性别决定系统的分化可能晚于杨、柳之间的物种分化, 性染色体在杨、柳这2个姊妹属中分别独立起源和进化, 演化形成XY和ZW两种完全不同的性别决定系统。我们对木本植物中已鉴别的性别决定基因进行了归类总结(表1)。
无花果(Ficus carica)属桑科(Moraceae)榕属(Ficus), 隐头花序, 根据其花类型和结实情况分为4种: 野生型(caprifig)、斯密尔那型(Smyrna)、圣比罗型(San Pedro)和普通型(common) (乔峰等, 2018)。野生型雌雄同花(由雄花和短柱雌花构成), 主要为无花果传粉蜂的寄主, 因此又被认为是无花果雄株; 而其它3种类型仅产生长柱雌花, 无花果的性别决定类型为XY型(Valdeyron and Lloyd, 1979)。Mori等(2017)构建了无花果基因组草图, 并结合家系连锁分析和全基因组关联分析(GWAS)将性别位点定位在1号染色体约100 kb的区间内。进一步分析该区间内所有SNP位点与性别的相关性, 发现有2个SNP位点(造成错义突变)与性别呈中等相关且在雄株中为杂合, 这2个SNP位点位于同一基因(RAN1)内部, 该基因编码铜转运ATP酶, 参与乙烯信号响应, 在雌株果实中表达量最高, 因此推测RAN1是无花果性别决定候选基因。
阿月浑子(Pistacia vera)又名开心果, 是漆树科(Anacardiaceae)黄连木属(Pistacia)的重要经济树种, 其性别早期鉴定具有较大的理论意义和应用价值.为开发与性别连锁的SNP位点用于分子标记辅助选择, Kafkas等(2015)随机挑选了阿月浑子杂交F1代中的8雄8雌, 与亲本分别进行RAD测序, 从筛选出的38个与性别连锁的SNP标记中开发出4对可以用于性别早期鉴定的引物, 利用这些引物对166份种质资源进行性别鉴定的准确率高达100%.此外, 38个与性别连锁的SNP位点均为雌株杂合、雄株纯合.该研究首次确定了阿月浑子的性别决定系统为ZW型. ... A Y-chromosome- encoded small RNA acts as a sex determinant in persimmons 2 2014 ... 柿属(Diospyros)植物不仅染色体倍性复杂, 性别类型也较为丰富, 存在雌株、雄株和雌雄异花同株等多种类型(Yonemori et al., 1993).近年来, 柿雌雄花性别分化的调控机理研究取得了重要进展.柿属二倍体植物君迁子(D. lotus)为雌雄异株, 成熟雄花3朵合生、呈聚伞状花序, 雌花1朵单生, 其性别决定系统为XY型(Akagi et al., 2013).在没有参考基因组序列的情况下, 研究人员对君迁子种内杂交F1代个体(32雌株和25雄株)分别进行基因组从头测序, 根据雌、雄性别将测序数据合并建成2个序列库, 通过对这2个序列库进行K-mer比较分析, 筛选出雄株特异的K-mer片段, 进行局部组装, 最终得到Y染色体上约1 Mb的雄株特异区间.结合雌雄花芽转录组差异表达分析, 结果表明该区间含有22个性别决定候选基因, 其中仅有1个基因(OGI)只在雄花中表达.此外, 通过差异表达分析还检测到1个位于常染色体上、仅在雌花中表达的基因(MeGI).序列分析显示, OGI基因能够转录形成发卡结构, 产生小RNA分子; MeGI基因编码HD-Zip蛋白(同源异型-亮氨酸拉链蛋白), 属于高等植物特有的转录因子, 且OGI产生的小RNA能够介导MeGI基因沉默, 这也是雄花中未检测到MeGI基因表达的原因.过表达MeGI基因的拟南芥(Arabidopsis thaliana)和烟草(Nicotiana tabacum)出现雄蕊缺损、变短和花粉萌发率降低等表型, 表明MeGI基因能够抑制雄蕊发育.由此证明, 位于Y染色体特异片段上的OGI基因决定君迁子的性别, 该基因通过产生小 RNA抑制MeGI基因在雄株中表达, 从而促进雄蕊发育; 而雌株由于没有OGI基因, MeGI基因能够正常表达, 导致雌花中的雄蕊退化(Akagi et al., 2014).随着君迁子雄株全基因组序列图谱的完成, OGI和MeGI基因的起源以及柿属植物性别决定系统的进化机制得到进一步明晰.Ks分析表明, 君迁子基因组在进化过程中经历了1次柿属植物特有的全基因组重复事件(Dd-α WGD), MeGI (Chr. 13)由其旁系同源基因SiMeGI (Chr. 4)经过全基因组重复或片段重复而来, MeGI和SiMeGI分化后演化出新的功能, 能够抑制雄蕊发育, 而SiMeGI仍保留祖先基因的功能, 调控花器官发育但不影响性别分化(Akagi et al., 2020).基于以上发现, Akagi等(2020)提出柿属植物从雌雄同株向雌雄异株的演化模型: 在柿子祖先基因组中仅有SiMeGI基因, 其经历Dd-α WGD产生MeGI, MeGI在演化过程中获得新的功能, 导致雌株出现; MeGI经过片段重复产生OGI, OGI反过来抑制MeGI的表达, 导致雄株产生.因此, 位于Y染色体特异片段上的OGI通过调控MeGI基因表达与否决定君迁子雌雄性别.MeGI是1个重要的整合器, 它通过整合影响下游靶基因的表达进而影响雄蕊或雌蕊的发育.综合利用雌雄花芽转录组差异表达和共表达网络, Yang等(2019)预测了MeGI基因下游调控网络.该基因通过KNOX/ OFP/GRF激活细胞分裂素/生长素/赤霉素途径相关基因促进雌蕊发育; 同时通过SVP/SOC抑制PI的表达来抑制雄蕊发育. ...
阿月浑子(Pistacia vera)又名开心果, 是漆树科(Anacardiaceae)黄连木属(Pistacia)的重要经济树种, 其性别早期鉴定具有较大的理论意义和应用价值.为开发与性别连锁的SNP位点用于分子标记辅助选择, Kafkas等(2015)随机挑选了阿月浑子杂交F1代中的8雄8雌, 与亲本分别进行RAD测序, 从筛选出的38个与性别连锁的SNP标记中开发出4对可以用于性别早期鉴定的引物, 利用这些引物对166份种质资源进行性别鉴定的准确率高达100%.此外, 38个与性别连锁的SNP位点均为雌株杂合、雄株纯合.该研究首次确定了阿月浑子的性别决定系统为ZW型. ... The persimmon genome reveals clues to the evolution of a lineage-specific sex determination system in plants 2 2020 ... 柿属(Diospyros)植物不仅染色体倍性复杂, 性别类型也较为丰富, 存在雌株、雄株和雌雄异花同株等多种类型(Yonemori et al., 1993).近年来, 柿雌雄花性别分化的调控机理研究取得了重要进展.柿属二倍体植物君迁子(D. lotus)为雌雄异株, 成熟雄花3朵合生、呈聚伞状花序, 雌花1朵单生, 其性别决定系统为XY型(Akagi et al., 2013).在没有参考基因组序列的情况下, 研究人员对君迁子种内杂交F1代个体(32雌株和25雄株)分别进行基因组从头测序, 根据雌、雄性别将测序数据合并建成2个序列库, 通过对这2个序列库进行K-mer比较分析, 筛选出雄株特异的K-mer片段, 进行局部组装, 最终得到Y染色体上约1 Mb的雄株特异区间.结合雌雄花芽转录组差异表达分析, 结果表明该区间含有22个性别决定候选基因, 其中仅有1个基因(OGI)只在雄花中表达.此外, 通过差异表达分析还检测到1个位于常染色体上、仅在雌花中表达的基因(MeGI).序列分析显示, OGI基因能够转录形成发卡结构, 产生小RNA分子; MeGI基因编码HD-Zip蛋白(同源异型-亮氨酸拉链蛋白), 属于高等植物特有的转录因子, 且OGI产生的小RNA能够介导MeGI基因沉默, 这也是雄花中未检测到MeGI基因表达的原因.过表达MeGI基因的拟南芥(Arabidopsis thaliana)和烟草(Nicotiana tabacum)出现雄蕊缺损、变短和花粉萌发率降低等表型, 表明MeGI基因能够抑制雄蕊发育.由此证明, 位于Y染色体特异片段上的OGI基因决定君迁子的性别, 该基因通过产生小 RNA抑制MeGI基因在雄株中表达, 从而促进雄蕊发育; 而雌株由于没有OGI基因, MeGI基因能够正常表达, 导致雌花中的雄蕊退化(Akagi et al., 2014).随着君迁子雄株全基因组序列图谱的完成, OGI和MeGI基因的起源以及柿属植物性别决定系统的进化机制得到进一步明晰.Ks分析表明, 君迁子基因组在进化过程中经历了1次柿属植物特有的全基因组重复事件(Dd-α WGD), MeGI (Chr. 13)由其旁系同源基因SiMeGI (Chr. 4)经过全基因组重复或片段重复而来, MeGI和SiMeGI分化后演化出新的功能, 能够抑制雄蕊发育, 而SiMeGI仍保留祖先基因的功能, 调控花器官发育但不影响性别分化(Akagi et al., 2020).基于以上发现, Akagi等(2020)提出柿属植物从雌雄同株向雌雄异株的演化模型: 在柿子祖先基因组中仅有SiMeGI基因, 其经历Dd-α WGD产生MeGI, MeGI在演化过程中获得新的功能, 导致雌株出现; MeGI经过片段重复产生OGI, OGI反过来抑制MeGI的表达, 导致雄株产生.因此, 位于Y染色体特异片段上的OGI通过调控MeGI基因表达与否决定君迁子雌雄性别.MeGI是1个重要的整合器, 它通过整合影响下游靶基因的表达进而影响雄蕊或雌蕊的发育.综合利用雌雄花芽转录组差异表达和共表达网络, Yang等(2019)预测了MeGI基因下游调控网络.该基因通过KNOX/ OFP/GRF激活细胞分裂素/生长素/赤霉素途径相关基因促进雌蕊发育; 同时通过SVP/SOC抑制PI的表达来抑制雄蕊发育. ...
阿月浑子(Pistacia vera)又名开心果, 是漆树科(Anacardiaceae)黄连木属(Pistacia)的重要经济树种, 其性别早期鉴定具有较大的理论意义和应用价值.为开发与性别连锁的SNP位点用于分子标记辅助选择, Kafkas等(2015)随机挑选了阿月浑子杂交F1代中的8雄8雌, 与亲本分别进行RAD测序, 从筛选出的38个与性别连锁的SNP标记中开发出4对可以用于性别早期鉴定的引物, 利用这些引物对166份种质资源进行性别鉴定的准确率高达100%.此外, 38个与性别连锁的SNP位点均为雌株杂合、雄株纯合.该研究首次确定了阿月浑子的性别决定系统为ZW型. ... The genome of black cottonwood, Populus trichocarpa (Torr. & Gray) 1 2006 ... 综上, 大部分杨树性别决定系统为XY型, 而大部分柳树性别决定系统为ZW型.比较基因组学显示, 杨树和柳树起源于共同的祖先, 基因组具有高度的共线性关系(Tuskan et al., 2006; Dai et al., 2014).但它们的性染色体分别由不同的常染色体进化而来: 杨树的性染色体是19号染色体, 而柳树的性染色体是15号染色体(Hou et al., 2015).细胞学研究表明, 杨树和柳树的性染色体均未出现形态上的差异, 尚处于进化早期阶段(Peto, 1938).由此可见, 性别决定系统的分化可能晚于杨、柳之间的物种分化, 性染色体在杨、柳这2个姊妹属中分别独立起源和进化, 演化形成XY和ZW两种完全不同的性别决定系统.我们对木本植物中已鉴别的性别决定基因进行了归类总结(表1). ... Genome sequence comparison reveals a candidate gene involved in male-hermaphrodite differentiation in papaya (Carica papaya) trees 1 2015 ... 番木瓜性别决定机制曾存在多种假说(Ming et al., 2007), 目前被普遍接受的是Horovitz和Jiménez (1967)根据属间杂交结果提出的XX-XY模型, 认为番木瓜雌株、雄株和两性株的基因型分别是XX、XY和XY2, Y2染色体由Y突变而来, Y和Y2染色体上均含有致死因子, 因此不存在YY、YY2和Y2Y2基因型的后代.由于雄株与两性株的Y染色体相似性很高(Liu et al., 2004), Ming等(2007)用Yh替代Y2, 用XYh表示两性株基因型.染色体序列比对和共线性分析显示, Y和Yh相似性高达99.60%.进一步分析发现Yh起源于Y染色体, 在约4 000年前完成分化, 在番木瓜长期的人工驯化过程中Yh被保留和固定下来(VanBuren et al., 2015).精细定位和FISH结果显示, 番木瓜性别决定基因位于LG1着丝粒附近约8 Mb的区间内, 该区间重组抑制严重, 且富含重复序列和转座子, 基因克隆难度较高(Yu et al., 2007; Wang et al., 2012).近年来, 基于转录组测序多个研究团队相继报道了番木瓜性别决定候选基因.Urasaki等(2012)通过转录组差异表达分析, 发现与序列标签Cp2671对应的基因编码1个MADS-box转录因子, 与拟南芥调控花期的转录因子SVP存在85%的序列相似性.SVP-like基因是Y和Yh染色体特异基因, 仅在雄花和两性株中表达, 由此提出SVP-like基因是番木瓜性别决定候选基因.通过比较MSY (male-specific region of Y)和HSY (hermaphrodite-specific region of Yh)的序列, 发现Y染色体上的SVP-like基因编码1个完整蛋白, 但Yh上SVP-like基因内有转座子插入, 编码不完整蛋白(Ueno et al., 2015).基于基因表达差异和选择性剪切(alternative splicing, AS)分析, Lee等(2018)同样认为CpSVPL (SVP-like)是番木瓜性别决定基因, 他们还提出另外2个性别相关基因CpSERK和CpCAF1AL.其中, CpSERK基因在不同性别花中表达量差异不显著, 但存在AS; CpCAF1AL在雄花和正常发育的两性花中表达量较高, 不存在AS, 在发育不正常的两性花中表达量较低且存在AS.Zerpa-Catanho等(2019)通过比较3种不同性别和不同发育时期番木瓜花RNA-seq数据, 发现CpMS1仅在花器官中表达, 且在雄花和两性花发育前期表达量较高, 该基因在拟南芥、大麦(Hordeum vulgare)和水稻(Oryza sativa)等植物中的同源基因功能相似, 能够调控花粉发育, 但由于CpMS1基因位于2号染色体, 且3种性别的番木瓜均含有该基因, 因此该基因不具有性别决定功能.最近, Zhou等(2019)通过对不同季节雌、雄花全基因组甲基化水平和转录组差异表达基因进行分析, 发现雄花基因组甲基化水平低于雌花, 常染色体上CpSVP和CpAP1基因的去甲基化可能是导致雄花比雌花早开放的原因.此外, 他们还发现CpARR5基因在雄花中的表达量和甲基化水平均较雌花低, 在检测到的10个与激素信号转导相关的差异表达基因中, CpARR5基因是唯一存在甲基化修饰位点差异的基因, 推测甲基化介导的植物激素稳态的改变可能影响番木瓜性别. ... Digital transcriptome analysis of putative sex-determination genes in papaya (Carica papaya) 2 2012 ... 番木瓜性别决定机制曾存在多种假说(Ming et al., 2007), 目前被普遍接受的是Horovitz和Jiménez (1967)根据属间杂交结果提出的XX-XY模型, 认为番木瓜雌株、雄株和两性株的基因型分别是XX、XY和XY2, Y2染色体由Y突变而来, Y和Y2染色体上均含有致死因子, 因此不存在YY、YY2和Y2Y2基因型的后代.由于雄株与两性株的Y染色体相似性很高(Liu et al., 2004), Ming等(2007)用Yh替代Y2, 用XYh表示两性株基因型.染色体序列比对和共线性分析显示, Y和Yh相似性高达99.60%.进一步分析发现Yh起源于Y染色体, 在约4 000年前完成分化, 在番木瓜长期的人工驯化过程中Yh被保留和固定下来(VanBuren et al., 2015).精细定位和FISH结果显示, 番木瓜性别决定基因位于LG1着丝粒附近约8 Mb的区间内, 该区间重组抑制严重, 且富含重复序列和转座子, 基因克隆难度较高(Yu et al., 2007; Wang et al., 2012).近年来, 基于转录组测序多个研究团队相继报道了番木瓜性别决定候选基因.Urasaki等(2012)通过转录组差异表达分析, 发现与序列标签Cp2671对应的基因编码1个MADS-box转录因子, 与拟南芥调控花期的转录因子SVP存在85%的序列相似性.SVP-like基因是Y和Yh染色体特异基因, 仅在雄花和两性株中表达, 由此提出SVP-like基因是番木瓜性别决定候选基因.通过比较MSY (male-specific region of Y)和HSY (hermaphrodite-specific region of Yh)的序列, 发现Y染色体上的SVP-like基因编码1个完整蛋白, 但Yh上SVP-like基因内有转座子插入, 编码不完整蛋白(Ueno et al., 2015).基于基因表达差异和选择性剪切(alternative splicing, AS)分析, Lee等(2018)同样认为CpSVPL (SVP-like)是番木瓜性别决定基因, 他们还提出另外2个性别相关基因CpSERK和CpCAF1AL.其中, CpSERK基因在不同性别花中表达量差异不显著, 但存在AS; CpCAF1AL在雄花和正常发育的两性花中表达量较高, 不存在AS, 在发育不正常的两性花中表达量较低且存在AS.Zerpa-Catanho等(2019)通过比较3种不同性别和不同发育时期番木瓜花RNA-seq数据, 发现CpMS1仅在花器官中表达, 且在雄花和两性花发育前期表达量较高, 该基因在拟南芥、大麦(Hordeum vulgare)和水稻(Oryza sativa)等植物中的同源基因功能相似, 能够调控花粉发育, 但由于CpMS1基因位于2号染色体, 且3种性别的番木瓜均含有该基因, 因此该基因不具有性别决定功能.最近, Zhou等(2019)通过对不同季节雌、雄花全基因组甲基化水平和转录组差异表达基因进行分析, 发现雄花基因组甲基化水平低于雌花, 常染色体上CpSVP和CpAP1基因的去甲基化可能是导致雄花比雌花早开放的原因.此外, 他们还发现CpARR5基因在雄花中的表达量和甲基化水平均较雌花低, 在检测到的10个与激素信号转导相关的差异表达基因中, CpARR5基因是唯一存在甲基化修饰位点差异的基因, 推测甲基化介导的植物激素稳态的改变可能影响番木瓜性别. ...
阿月浑子(Pistacia vera)又名开心果, 是漆树科(Anacardiaceae)黄连木属(Pistacia)的重要经济树种, 其性别早期鉴定具有较大的理论意义和应用价值.为开发与性别连锁的SNP位点用于分子标记辅助选择, Kafkas等(2015)随机挑选了阿月浑子杂交F1代中的8雄8雌, 与亲本分别进行RAD测序, 从筛选出的38个与性别连锁的SNP标记中开发出4对可以用于性别早期鉴定的引物, 利用这些引物对166份种质资源进行性别鉴定的准确率高达100%.此外, 38个与性别连锁的SNP位点均为雌株杂合、雄株纯合.该研究首次确定了阿月浑子的性别决定系统为ZW型. ... Sex differences and flowering phenology in the common fig, Ficus carica L 1 1979 ... 无花果(Ficus carica)属桑科(Moraceae)榕属(Ficus), 隐头花序, 根据其花类型和结实情况分为4种: 野生型(caprifig)、斯密尔那型(Smyrna)、圣比罗型(San Pedro)和普通型(common) (乔峰等, 2018).野生型雌雄同花(由雄花和短柱雌花构成), 主要为无花果传粉蜂的寄主, 因此又被认为是无花果雄株; 而其它3种类型仅产生长柱雌花, 无花果的性别决定类型为XY型(Valdeyron and Lloyd, 1979).Mori等(2017)构建了无花果基因组草图, 并结合家系连锁分析和全基因组关联分析(GWAS)将性别位点定位在1号染色体约100 kb的区间内.进一步分析该区间内所有SNP位点与性别的相关性, 发现有2个SNP位点(造成错义突变)与性别呈中等相关且在雄株中为杂合, 这2个SNP位点位于同一基因(RAN1)内部, 该基因编码铜转运ATP酶, 参与乙烯信号响应, 在雌株果实中表达量最高, 因此推测RAN1是无花果性别决定候选基因. ... Origin and domestication of papaya Yh chromosome 2 2015 ... 番木瓜科(Caricaceae)大多是雌雄异株植物.番木瓜(Carica papaya)是其中仅有的2种单全异株(trioecious)植物之一, 其种群内既有雌株和雄株, 还具有两性株(雌雄同花), 雄花中可见心皮遗迹, 而雌花中没有雄蕊痕迹, 在不良环境条件下, 雄株和两性株可以发生性别转换(Aryal and Ming, 2014).番木瓜由于生长周期短、性别多样且有良好的遗传学和基因组学研究基础, 被作为植物性别决定研究的模式植物(Ming et al., 2007), 特别是对其性染色体分化的系列研究(Liu et al., 2004; Yu et al., 2008a, 2008b; Ming et al., 2011; Wang et al., 2012; VanBuren et al., 2015), 为人们认识和理解植物性染色体起源与进化提供了非常重要的证据. ...
阿月浑子(Pistacia vera)又名开心果, 是漆树科(Anacardiaceae)黄连木属(Pistacia)的重要经济树种, 其性别早期鉴定具有较大的理论意义和应用价值.为开发与性别连锁的SNP位点用于分子标记辅助选择, Kafkas等(2015)随机挑选了阿月浑子杂交F1代中的8雄8雌, 与亲本分别进行RAD测序, 从筛选出的38个与性别连锁的SNP标记中开发出4对可以用于性别早期鉴定的引物, 利用这些引物对166份种质资源进行性别鉴定的准确率高达100%.此外, 38个与性别连锁的SNP位点均为雌株杂合、雄株纯合.该研究首次确定了阿月浑子的性别决定系统为ZW型. ... Gene networks orchestrated by MeGI: a single-factor mechanism underlying sex determination in persimmon 1 2019 ... 柿属(Diospyros)植物不仅染色体倍性复杂, 性别类型也较为丰富, 存在雌株、雄株和雌雄异花同株等多种类型(Yonemori et al., 1993).近年来, 柿雌雄花性别分化的调控机理研究取得了重要进展.柿属二倍体植物君迁子(D. lotus)为雌雄异株, 成熟雄花3朵合生、呈聚伞状花序, 雌花1朵单生, 其性别决定系统为XY型(Akagi et al., 2013).在没有参考基因组序列的情况下, 研究人员对君迁子种内杂交F1代个体(32雌株和25雄株)分别进行基因组从头测序, 根据雌、雄性别将测序数据合并建成2个序列库, 通过对这2个序列库进行K-mer比较分析, 筛选出雄株特异的K-mer片段, 进行局部组装, 最终得到Y染色体上约1 Mb的雄株特异区间.结合雌雄花芽转录组差异表达分析, 结果表明该区间含有22个性别决定候选基因, 其中仅有1个基因(OGI)只在雄花中表达.此外, 通过差异表达分析还检测到1个位于常染色体上、仅在雌花中表达的基因(MeGI).序列分析显示, OGI基因能够转录形成发卡结构, 产生小RNA分子; MeGI基因编码HD-Zip蛋白(同源异型-亮氨酸拉链蛋白), 属于高等植物特有的转录因子, 且OGI产生的小RNA能够介导MeGI基因沉默, 这也是雄花中未检测到MeGI基因表达的原因.过表达MeGI基因的拟南芥(Arabidopsis thaliana)和烟草(Nicotiana tabacum)出现雄蕊缺损、变短和花粉萌发率降低等表型, 表明MeGI基因能够抑制雄蕊发育.由此证明, 位于Y染色体特异片段上的OGI基因决定君迁子的性别, 该基因通过产生小 RNA抑制MeGI基因在雄株中表达, 从而促进雄蕊发育; 而雌株由于没有OGI基因, MeGI基因能够正常表达, 导致雌花中的雄蕊退化(Akagi et al., 2014).随着君迁子雄株全基因组序列图谱的完成, OGI和MeGI基因的起源以及柿属植物性别决定系统的进化机制得到进一步明晰.Ks分析表明, 君迁子基因组在进化过程中经历了1次柿属植物特有的全基因组重复事件(Dd-α WGD), MeGI (Chr. 13)由其旁系同源基因SiMeGI (Chr. 4)经过全基因组重复或片段重复而来, MeGI和SiMeGI分化后演化出新的功能, 能够抑制雄蕊发育, 而SiMeGI仍保留祖先基因的功能, 调控花器官发育但不影响性别分化(Akagi et al., 2020).基于以上发现, Akagi等(2020)提出柿属植物从雌雄同株向雌雄异株的演化模型: 在柿子祖先基因组中仅有SiMeGI基因, 其经历Dd-α WGD产生MeGI, MeGI在演化过程中获得新的功能, 导致雌株出现; MeGI经过片段重复产生OGI, OGI反过来抑制MeGI的表达, 导致雄株产生.因此, 位于Y染色体特异片段上的OGI通过调控MeGI基因表达与否决定君迁子雌雄性别.MeGI是1个重要的整合器, 它通过整合影响下游靶基因的表达进而影响雄蕊或雌蕊的发育.综合利用雌雄花芽转录组差异表达和共表达网络, Yang等(2019)预测了MeGI基因下游调控网络.该基因通过KNOX/ OFP/GRF激活细胞分裂素/生长素/赤霉素途径相关基因促进雌蕊发育; 同时通过SVP/SOC抑制PI的表达来抑制雄蕊发育. ... A general model to explain repeated turnovers of sex determination in the Salicaceae 7 2020 ... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
本文的其它图/表
-
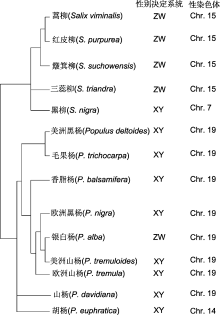 图1
已确定性别决定系统的杨柳科植物谱系树
图1
已确定性别决定系统的杨柳科植物谱系树
文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020)
|

 首页
首页