
 首页
首页

丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017)。植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分。人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019)。尽管如此, 仍有学者发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018)。这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循。当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020)。这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020)。尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义。
丁香属(Syringa)为木犀科落叶灌木或小乔木, 分布在亚洲温带地区及欧洲东南部(崔洪霞等, 2004)。全属有野生分布的原种和变种约30种(McKelvey, 1928; Rehder, 1940; 张美珍等, 1992)。中国是丁香属植物的自然分布中心, 约有18种(张美珍等, 1992)。丁香因花开繁盛馨香和抗逆性突出, 成为世界民众喜爱的经典观赏植物(藏淑英和李容辉, 1992), 也因含有丰富的抗氧化、抗炎症和抗心肌缺血等功能性次生代谢成分而成为中国和欧洲古老的民族药材(王宇希, 2013; 苏国柱等, 2015; Filipek et al., 2019)。我们通过检索获得了1998-2020年丁香属次生代谢相关文献80余篇, 剔除了因技术条件限制而使成分鉴定存疑的早期文献后, 选取58篇文献进行化合物汇总。文献涉及11个原种和变种及1个种间杂交种, 全属种质覆盖率为40%, 覆盖属下全部5个组系, 因而具有充分的系统学代表性。对丁香属众多已知的次生代谢产物进行整理, 并结合系统学背景及环境适应进行分析, 有助于在属级分类单元上理解次生代谢产物的分布格局, 为次生代谢调控的系统性研究提供新的启示, 可引导专属功能性成分的高效开发, 也为资源的系统性收集注入了源动力。
1 丁香属次生代谢产物及主要代谢途径
丁香属植物含有丰富的次生代谢产物, 具有相同母核的同类成分常因取代基团类型、数量、位置和空间构象的不同而数量众多。经整理我们发现已报道的次生代谢产物共有377个(附表1), 按其母核结构可分为10类: 木脂素类成分74个, 环/裂环烯醚萜类成分118个, 倍半萜类成分67个, 三萜类成分34个, 单萜类成分3个, 黄酮类成分13个, 苯丙素类成分59个, 脂肪酸类3个, 醌类2个, 生物碱类4个。其中含量较高的成分多是特定功能的重要贡献者, 其对应的代谢途径也是我们分析的重点。由于针对属内组系水平的化学多样性分析的客观性取决于特定物种的研究案例数量和属下分类等级上的物种覆盖度, 我们选取的文献所述物种在丁香属内涉及5个组系(表1)。在这些文献中, 中国原产的单种系羽叶丁香系(Ser. Pinnatifoliae) 21篇; 东亚和欧洲分布的欧丁香系(Ser. Syringa) 20篇, 其中东亚广布种紫丁香(S. oblata)和朝阳丁香(S. oblata var. dilatata)分别为10篇和3篇, 东南欧洲广布种欧洲丁香(S. vulgaris) 6篇, 中国西北-中西亚分布种阿富汗丁香(S. afghanica) 1篇; 东亚及远东广布的短花冠管组(Sect. Ligustrina)中的暴马丁香(S. amurensis) 6篇和日本丁香(S. reticulata) 3篇; 中国华北和东北分布的巧玲花系(Ser. Pubescentes)共6篇, 其中包括华北分布种巧玲花(S. pubescens)和东北分布种关东丁香(S. velutina)各3篇; 中国西南和东北亚分布的红丁香系(Ser. Villosae) 2篇, 其中包括西蜀丁香(S. komarowii)和辽东丁香(S. wolfii)各1篇。我们分别在组系(表2)和物种(表3) 2个分类水平上对次生代谢产物进行归类整理。通过python+seaborn软件包(
表1 已报道次生代谢成分的丁香属种质地理分布
Table 1
| 组系 | 物种 | 自然分布 | 海拔(m) | 经度(E) | 纬度(N) | 生态幅 |
|---|---|---|---|---|---|---|
| 欧丁香系 (Ser. Syringa) | 欧洲丁香(S. vulgaris) | 东南欧(阿尔卑斯山和喀尔巴阡山) | 1000-1200 | 5°36'-66°10' | 36°00'-67°46' | 广布种 |
| 紫丁香(S. oblata) | 中国东北、华北、西北、华东和川西北 | 300-2400 | 96°23'-135°02' | 34°19'-55°33' | 广布种 | |
| 朝阳丁香(S. oblata var. dilatata) | 中国华北-东北-朝鲜半岛 | 300-2400 | 40°51'-129°40' | 33°56'-53°19' | 广布种 | |
| 阿富汗丁香(S. afghanica) | 中国青海-阿富汗 | 60°29'-74°53' | 29°21'-38°27' | 局域种 | ||
| 羽叶丁香系 (Ser. Pinnatifoliae) | 羽叶丁香(S. pinnatifolia) | 贺兰山、陕西南部、甘肃、青海东部和四川南部 | 1700-3100 | 105°13'-112°32' | 26°03'-37°09' | 局域种 |
| 短花冠管组 (Sect. Ligustrina) | 暴马丁香(S. amurensis) | 中国东北和西北, 俄罗斯远东地区, 朝鲜 | 100-1200 | 103°04'-163°19' | 31°09'-72°33' | 广布种 |
| 日本丁香(S. reticulata) | 日本北部(北海道地区) | 139°20'-148°53' | 40°33'-45°33' | 局域种 | ||
| 巧玲花系 (Ser. Pubescentes) | 巧玲花(S. pubescens) | 河北、陕西东部、山西东部和河南 | 900-2100 | 109°29'-119°53' | 31°23'-42°37' | 局域种 |
| 关东丁香(S. velutina) | 辽宁和吉林长白山区 | 300-1200 | 118°53'-135°05' | 38°43'-53°33' | 局域种 | |
| 红丁香系 (Ser. Villosae) | 西蜀丁香(S. komarowii) | 甘肃南部、陕西南部、 四川和云南北部 | 105°31'-114°11' | 24°20'-35°28' | 局域种 | |
| 辽东丁香(S. wolfii) | 中国东北, 朝鲜 | 500-1600 | 118°53'-128°28' | 37°35'-72°33' | 局域种 |
表中仅列次生代谢成分已报道的9个原种和2个变种, 涉及丁香属下5个组系的所有分类等级。生态幅指丁香属内特定物种的地理分布跨度, 即其分布的经度、纬度或海拔数据中最小值与最大值之间的范围。广布种指经度跨度大于30°或纬度跨度大于20°的物种, 局域种指经度跨度小于30°或纬度跨度小于20°的物种。
The species with secondary metabolites reported in the table include nine native species and two varieties, involving four series and one section of Syringa. Ecological amplitude refers to the geographical span of a species, the range between the highest and lowest points of longitude, latitude and altitude. Widespread species refers to the species whose longitude span is more than 30° or latitude span is more than 20°, and local species refers to the species whose longitude span is less than 30° or latitude span is less than 20°.
表2 丁香属组系水平的次生代谢产物类别及各类成分计数(单位: 个)
Table 2
| 代谢产物类别 | 欧丁香系 (Ser. Syringa) | 羽叶丁香系 (Ser. Pinnatifoliae) | 短花冠管组 (Sect. Ligustrina) | 巧玲花系 (Ser. Pubescentes) | 红丁香系 (Ser. Villosae) |
|---|---|---|---|---|---|
| 环/裂环烯醚萜 | 96 | 6 | 17 | 9 | 2 |
| 倍半萜 | 8 | 46 | 6 | 10 | - |
| 苯丙素 | 30 | 5 | 11 | 8 | 8 |
| 木脂素 | 5 | 58 | 8 | 1 | 5 |
| 黄酮 | 8 | - | - | 5 | - |
| 三萜 | 15 | 8 | 2 | 2 | 12 |
| 单萜 | 3 | - | - | - | - |
| 脂肪酸 | 3 | - | - | - | - |
| 醌类 | - | 2 | - | - | - |
| 生物碱 | - | - | - | 1 | 3 |
| 合计 | 168 | 125 | 44 | 36 | 30 |
- 表示该类产物在特定组系中未见报道。成分来自根、茎、花、叶和茎皮多个器官。各类产物计数指具有相同母核但取代基团、基团数量、基团位置和空间构象不同的化合物个数。
- indicates that the components have not been reported in the series (section). The components came from multiple organs, including root, stem, flower, leaf and stem bark. The component counting refers to the number of the components with the same parent nucleus but different in substituent groups, group numbers, group positions and spatial conformations.
表3 丁香属物种水平的次生代谢产物类别及成分计数(单位: 个)
Table 3
| 代谢产物类别 | 欧丁香系 (Ser. Syringa) | 羽叶丁香系 (Ser. Pinna tifoliae) | 短花冠管组 (Sect. Ligustrina) | 巧玲花系 (Ser. Pubescentes) | 红丁香系 (Ser. Villosae) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 欧洲丁香 (S. vulgaris) | 紫丁香 (S. ob- lata) | 朝阳丁香 (S. oblata var. dilatata) | 阿富汗丁香 (S. afghanica) | 羽叶丁香 (S. pinnatifolia) | 暴马丁香 (S. amu- rensis) | 日本丁香 (S. reticulata) | 关东丁香 (S. velutina) | 巧玲花 (S. pubescens) | 辽东丁香 (S.wolfii) | 西蜀丁香 (S. komarowii) | |
| 环/裂环烯醚萜 | 60 | 16 | 10 | 20 | 6 | 13 | 5 | 7 | 3 | - | 2 |
| 倍半萜 | - | 6 | 2 | - | 46 | 6 | - | - | 10 | - | - |
| 苯丙素 | 28 | 3 | - | - | 5 | 6 | 5 | 7 | 1 | - | 8 |
| 木脂素 | 5 | - | - | - | 58 | 1 | 7 | 1 | - | - | 5 |
| 黄酮 | 5 | 5 | - | - | - | - | - | 5 | - | - | - |
| 三萜 | - | 14 | 2 | - | 8 | 2 | - | 2 | - | 2 | 10 |
| 单萜 | - | 3 | - | - | - | - | - | - | - | - | - |
| 脂肪酸 | 1 | 2 | - | - | - | - | - | - | - | - | - |
| 醌类 | - | - | - | - | 2 | - | - | - | - | - | - |
| 生物碱 | - | - | - | - | - | - | - | 1 | - | - | 3 |
| 合计 | 99 | 49 | 14 | 20 | 125 | 28 | 17 | 23 | 14 | 2 | 28 |
- 表示该类产物在特定物种中未见报道。成分来自根、茎、花、叶和茎皮。各类成分计数包括具有相同母核但取代基团、基团数量、基团位置和空间构象不同的化合物。阿富汗丁香、辽东丁香和西蜀丁香数据分别仅来源于1篇文献。
- indicate that the components have not been reported in correspoding species. The components came from root, stem, flower, leaf and stem bark. The component counting includes different compounds with the same parent nucleus but different in substituent groups, group numbers, group positions and spatial conformations. The data of S. afghanica, S. wolfii and S. komarowii were only obtained from one reference, respectively.
图1
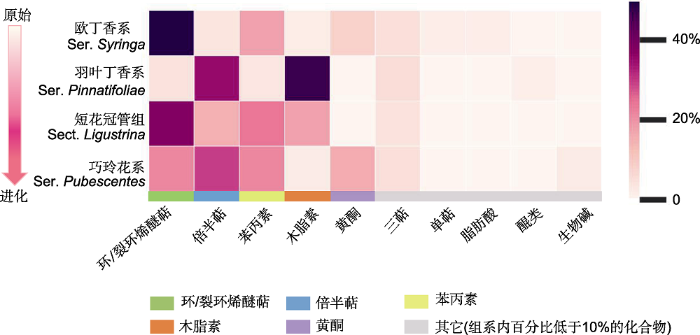
图1
丁香属组系水平的次生代谢成分相对占比
色块内颜色从深到浅表示各类产物的相对数量占比从高到低的连续变化(组系内特定类别产物的相对数量占比=特定类别产物的成分计数/该组系内所有类别产物计数总和), 同一类别产物包含具有相同母核但取代基团、基团数量、基团位置和空间构象不同的化合物。热图下方的不同颜色条表示不同类别产物。为保证成分计数占比分析的客观性, 图中每个物种收入的成分信息需至少来自3篇文献。欧丁香系的阿富汗丁香及红丁香系的辽东丁香和西蜀丁香因各仅有1篇文献而暂未收入。
Figure 1
Proportion of secondary metabolites in Syringa at the level of series (section)
The color in the block progressively varying from dark to light represents the continuous change in the percentage of components in the series (section) from high to low (the percentage of specific type of components in a series (section) = the number of specific type of components in a series (section)/the total number of components in this series (section)). The products of the same type include different components with the same parent nucleus but different in substituent groups, group numbers, group positions and conformations. The different color-stipes below the heatmap represent different types of products. In order to ensure the objective counting of the components, at least three reference were required for the metabolite information of each species. Therefore, the S. afghanica in Ser. Syringa as well as the S. wolfii and S. komarowii in Ser. Villosae has not been included because only one related reference was found.
图2
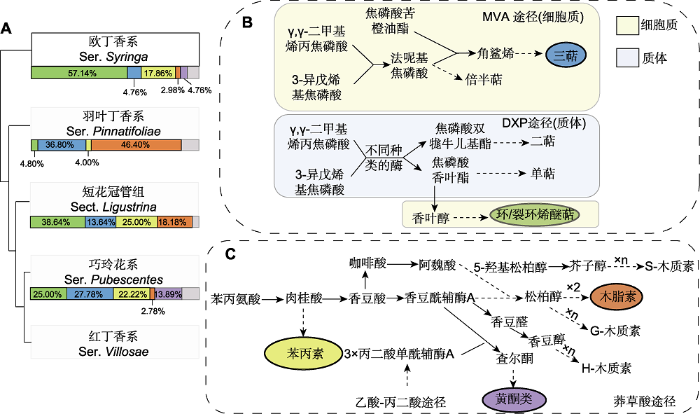
图2
丁香属内组系的系统演化关系和组系中各类产物的占比及其主要代谢途径
(A) 丁香属组系的系统演化关系(Li et al., 2012)及各类产物在各组系中的占比; (B) 甲戊二羟酸途径(MVA)和脱氧木酮糖磷酸酯途径(DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) 莽草酸途径(刘津等, 2016; 张旭等, 2019)。图(A)组系下方的色条表示各类产物(颜色含义同
Figure 2
The phylogenetic relationship of Syringa, and the percentage of metabolites and their main metabolic pathways at the level of series (section)
(A) The phylogenetic relationship (Li et al., 2012) and percentage of metabolites of Syringa at the level of series (section); (B) Mevalonic acid pathway (MVA) and Deoxyxylulose-5-P pathway (DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) Shikimic acid pathway (Liu et al., 2016; Zhang et al., 2019, in Chinese). The color-stripes below the series (section) indicate different types of metabolites in (A), and the percentage in the color-stripe represents the percentage of a given type of compounds within the series (section) (please see Figure 1 for details). The meanings of the color frames indicating the compound in (B) and (C) are the same as those of (A). The dotted lines in (B) and (C) represent the process of multi-step reactions.
萜类是丁香属中一类重要的次生代谢产物。一般而言, 萜类成分的合成涉及甲戊二羟酸途径和脱氧木酮糖途径。MVA途径在细胞质中进行, 主要合成倍半萜和三萜; DXP途径在质体中进行, 主要合成单萜、二萜和多萜等成分(图2B) (Mint Evolutionary Genomics Consortium, 2018)。从属下分类等级看, 欧丁香系占比高达57.14%的成分为环/裂环烯醚萜类化合物。在质体中, 3-异戊烯基焦磷酸(isopentenyl pyrophosphate, IPP)及其异构体γ,γ-二甲基烯丙焦磷酸(dimethylallyl diphosphate, DMAPP)头尾缩合生成单萜类及环/裂环烯醚萜的前体焦磷酸香叶酯(geranyl pyrophosphate, GPP) (董娟娥等, 2009)。相对于仅经过1次酶促反应就可使GPP生成单萜来说, 作为DXP途径的下游产物, 环/裂环烯醚萜的合成则要从质体输出到细胞质后再经历一系列酶的催化才能形成(图2B) (杨然等, 2018)。对于干旱半干旱地区局域分布的羽叶丁香而言, 同样以IPP和DMAPP为底物生成的倍半萜类(36.80%) (图2B)主要通过MVA途径在细胞质中产生, 而环/裂环烯醚萜的合成则大量减少。短花冠管组的倍半萜有所保留, 但其占比的优势度下降至13.64%, 环/裂环烯醚萜类占比升至38.64%。巧玲花系萜类成分特征与短花冠管组相似, 同时保留了倍半萜(27.78%)与环/裂环烯醚萜(25.62%)两类优势化合物。
莽草酸途径是芳香族化合物的生成途径。丁香属中的苯丙素类、木脂素类和黄酮类成分都经由莽草酸途径合成, 只是在代谢路径长度上各有不同(图2C)。同样从属下分类等级看, 经过较短的莽草酸途径形成的苯丙素类产物在欧丁香系中有较为优势的表达, 使该系产物占比达到全部产物的17.86%。该途径在羽叶丁香中则一直延长至下游产物木脂素类的生成, 其占比高达46.40%, 而上游产物苯丙素类成分仅在羽叶丁香的所有成分中占4.00%。在短花冠管组中苯丙素类及木脂素类化合物含量均超过10%。该组中下游的木脂素占比优势相对于干旱环境中的羽叶丁香有所下调, 但上游的苯丙素类成分呈现一定丰度, 两类产物加和后占该组化合物的43.18%。巧玲花系中苯丙氨酸向相对复杂的黄酮合成方向延伸, 其含量升至13.89%。黄酮类化合物的合成需要结合乙酸-丙二酸途径, 这表明需经过更长的代谢途径和更加复杂的合成过程(图2C)。
2 丁香属次生代谢产物与系统演化及环境的关联
对于丁香属内次生代谢产物的讨论离不开物种分类和演化的系统学背景。从经典分类以及既往分子生物学的研究结果看, 丁香属系统演化关系基本明确, 其结果差异主要在于在先分化类群的不同(张美珍等, 1992; Kim and Jansen, 1998; Li et al., 2012)。因此,我们依据欧洲丁香与紫丁香存在欧亚间断分布的事实(Kim and Jansen, 1998), 以及核DNA分析在物种演化关系判定上的依据(Li et al., 2012), 支持灌木型欧丁香系是丁香属最先分化的类群。羽叶丁香系在欧丁香系之后独立分化, 乔木型短花冠管组位居灌木型组系之间为中间分化类群(Li et al., 2012), 而对于巧玲花系和红丁香系是丁香属内后续分化类群的观点则不存异议。据此, 我们在进行系统关联讨论时采用Li等(2012)所报道的系统演化模式。
2.1 丁香属次生代谢产物在演化梯度上的优势变化
本研究表明, 丁香属在组系分化的先后梯度上表现为成分的优势度降低而化学多样性水平升高。在先分化的组系有更具优势的特定类型代谢产物, 而后续分化的组系特定类型成分的优势度降低而多样性水平升高(图1)。首先分化的欧丁香系物种虽有苯丙素、木脂素、黄酮、单萜、倍半萜、三萜和脂肪酸类等众多类型的代谢产物, 但丝毫不影响环/裂环烯醚萜类产物的绝对占比优势(高达57.14%)。这使得紫丁香和欧洲丁香能在更多样的环境选择压力下生存, 包括由经纬度和海拔的较大变幅所造成的气温、光照以及土壤环境的巨大差异, 欧丁香系中的欧洲丁香能够在巴尔干半岛南部山区中广泛分布, 紫丁香也能在中国北方的广阔区域中繁衍生息(表1, 表3)。对于较先分化的羽叶丁香系, 相对占比高达46.40%的木脂素同样具有优势。在此后持续的演化分异中, 特定产物的优势出现降低趋势。如果说处在进化中间位置的短花冠管组尚有37.64%的环/裂环烯醚萜类成分占比, 相对于较先分化的羽叶丁香系已经明显降低的话, 那么后续分化的巧玲花系很难看出特定化合物的绝对优势, 其中环/裂环烯醚萜类(25.00%)、倍半萜类(27.78%)和苯丙素类(22.22%)成分的数量占比则更为均衡(图1)。
2.2 丁香属次生代谢的系统学保守性
丁香属中的环/裂环烯醚萜类和苯丙素类产物呈现较强的系统遗传保守性, 甚至遥远的地理隔离也未造成近缘种优势代谢产物的明显差异。紫丁香和欧洲丁香是系统学意义上的姊妹系(Li et al., 2012), 虽然二者在自然分布上存在着东亚与巴尔干半岛的遥远亚欧间断分布(表1), 却均具有丰富的环/裂环烯醚萜类和苯丙素类产物(表3)。苯丙素类产物形成的保守性还体现在短花冠管组的2个近缘种暴马丁香和日本丁香中。暴马丁香广泛分布于中国北方及远东地区, 日本丁香仅在日本呈局域分布, 但2个系统学的近缘种同样具有占比相近的苯丙素类成分(表1, 表3)。由此表明, 环/裂环烯醚萜类和苯丙素类成分的形成具有较强的遗传保守性, 强烈的保守性没有因遥远的地理距离而消弱。与此同时, 在相邻的近缘组系中出现了较高占比的特定类别化合物的连续分布(图1)。图2A显示, 木脂素以较高的占比出现在相邻的羽叶丁香系(46.40%)和短花冠管组(18.18%)中; 倍半萜分别以36.80%、13.64%和27.78%的较高占比出现在相邻的羽叶丁香系、短花冠管组和巧玲花系中; 苯丙素以25.00%和22.22%的占比出现在相邻的短花冠管组和巧玲花系中; 环/裂环烯醚萜类分别以38.64%和25.00%的较高占比出现在相邻的短花冠管组和巧玲花系中。值得一提的是, 在针对同一个物种的诸多文献进行化合物汇总时, 我们发现欧洲丁香的样品取自罗马尼亚(Varga et al., 2019)和波兰(Filipek et al., 2019)多地, 但这些来自不同环境的同一物种却具有高度相似的次生代谢产物。这表明在丁香属特定的次生代谢途径中确实存在无法被环境异质性改变的系统学保守性。
2.3 从代谢角度看丁香属系统演化与地理分布跨度的对应关系
在分别从系统演化和地理环境角度讨论丁香属次生代谢产物的分布趋势之后, 需要面对的问题还有代谢背景下的系统演化与地理分布跨度的关系。研究表明, 紫丁香和欧洲丁香是丁香属系统学原始组系中的2个近缘姊妹系物种(Li et al., 2012), 它们也分别是东亚和欧洲的广布种(张美珍等, 1992; Fiala, 2008), 而后续分化的红丁香系(Li et al., 2012)中的大多数物种都属于生态幅狭窄的局域种。已知温度和水分是限制物种扩散的主要环境因子(高坤等, 2018; 魏华等, 2018), 丁香地理分布范围的差异一定程度上由对温度和水分敏感的光合同化产物形成和水分利用效率等初生代谢机制的差别决定(Cui et al., 2016)。紫丁香和欧洲丁香的广布表明其初生代谢机制保证了它们能够适应东亚和东南欧洲的气候变化, 同时强大的系统学保守性使相距万里的2个系统学意义上的近缘姊妹种均具有环/裂环烯醚萜和苯丙素类产物(表1, 表3), 使它们有能力通过这两类优势次生代谢产物的形成, 增强对小环境中生物及非生物扰动的适应能力, 从而改善植株的生长状况并实现高效的繁衍生息(Konno et al., 1999; 刘盟盟等, 2017; Sharma et al., 2019)。而对于性喜冷凉的红丁香系中的红丁香和匈牙利丁香2个近缘种, 气孔较大且密度较低的叶片表皮特征很可能使其在初生代谢热量耗散和叶片水分保持能力上存在限制(高艳等, 2008), 因此其分布局限在高海拔的冷凉或邻水生境(张美珍等, 1992; Lendvay et al., 2016)而成为地理分布上的局域种。在这类水热条件适宜的小环境中, 这些较晚分化的局域种不必应对地域扩散中温度和水分的巨大变化, 有条件形成更加多样的次生代谢产物, 在适宜的生境中最大限度地改善生长质量。这种“初生代谢决定生存格局, 次生代谢改善生活质量”(刘晓侠等, 2015)的情况也发生在同样进化却局域分布的巧玲花系中(图1)。
2.4 极端环境驱动特定代谢产物的优势表达
极端环境压力可能会使特定的代谢路径延长。木脂素是丁香属中独具特色的成分, 现有文献显示其在羽叶丁香系和短花冠管组中占据优势。在单种系的羽叶丁香中木脂素占比高达46.40%, 这与其生长的极端干旱环境紧密相关。羽叶丁香生长在贺兰山东、西山麓以及陕西南部、甘肃和青海, 该地区原生境大多降水稀少, 巨大的水分胁迫压力使羽叶丁香的地下生物量投入极大, 以粗壮的根系实现土壤深层水分的向性生长(Cui et al., 2016; 高坤等, 2018)。莽草酸途径化合物的积累通常有助于增强植物对水分胁迫的耐受性(刘盟盟等, 2017; Sharma et al., 2019), 而该途径中形成的木脂素可在木质部木质化后成为一种后注入型树脂, 使植物在水分胁迫而韧皮部运输减少的情况下更具干旱耐受性(Gaylord et al., 2013; Zhang et al., 2018)。这种被极端干旱强化的木脂素代谢途径是以牺牲莽草酸途径中大量上游产物苯丙素为代价以保证下游产物木脂素的产生, 助力粗壮根系的形成以实现土壤深层的趋向性生长, 从而最大程度地吸收生存所需水分。与此同时, 吸引传粉者也是其需要兼顾的因素, 在1 700-3 100 m的高山生境中, 传粉昆虫比低海拔地区更为稀少, 这使得羽叶丁香在萜类途径中更多地利用底物焦磷酸二甲烯丙酯和焦磷酸烯丙酯向法尼基焦磷酸合成方向延伸, 成为丁香属以倍半萜为优势萜类成分的组系。利用较短途径获得的大量倍半萜可在一定程度上实现对传粉昆虫的诱集(Chadwick et al., 2013)。
3 研究展望
我们对丁香属已知次生代谢产物在组系水平的表达进行了归纳, 并对其系统学演化关联信息进行了挖掘。文献涉及的物种实现了全属组系等级的全覆盖, 提示化学多样性的表象背后存在着系统学的深刻影响。丁香属内苯丙素类和环/裂环烯醚萜类产物具有系统保守性, 启示研究者可进一步整合基因组和代谢组学的方法, 在高度覆盖全属种质的前提下, 从众多植物属下的组系甚至种水平上更为深入地揭示代谢调控因子的系统演化机制, 同时为更多类群功能性成分的高效开发提供启示和引导, 为资源的系统性收集注入源动力。
附表1 丁香属已发表的次生代谢产物信息汇总
Appendix table 1 Summary of published secondary metabolites of Syringa
参考文献

为了探讨放牧过程中冷蒿(Artemisia frigida)酚酸及其抗氧化酶活性对牲畜采食和践踏损伤的响应, 该研究对盆栽冷蒿幼苗枝叶采用不同程度(轻度、中度和重度)机械损伤的方式模拟放牧强度, 利用高效液相色谱技术测定了机械损伤处理6 h后冷蒿枝叶和根系9种酚酸含量的变化; 同时对多酚氧化酶(PPO)、苯丙氨酸氨裂合酶(PAL)和抗氧化酶活性进行了测定, 分析了酚酸含量与PPO和PAL活性之间的相关性。结果表明: 轻度机械损伤可以诱导冷蒿枝叶和根系中PPO、PAL和抗氧化酶迅速做出响应, 活性显著升高; 随着机械损伤强度增加, 冷蒿枝叶游离态咖啡酸、丁香酸、阿魏酸和肉桂酸含量显著增加, 与对照相比中度损伤处理分别增加了150.4%、93.5%、154.4%和121.7%, 与PAL活性呈正相关关系; 游离态绿原酸和邻苯二酚含量减少, 与对照相比重度损伤处理后降低了91.1%和69.3%, 与PPO活性呈负相关关系; 与对照相比重度损伤处理下没食子酸和原儿茶酸分别增加了280.6%和215.7%。随损伤强度增加, 根系中9种游离态酚酸含量均显著增加, 其含量均小于叶片。机械损伤导致冷蒿枝叶中游离态和结合态酚酸总量增加, 根系中结合态酚酸减少。上述结果表明机械损伤首先诱导冷蒿体内应激防御系统中抗氧化酶和酚酸合成关键酶活性升高, 然后促进抗氧化物质酚酸的积累, 调节冷蒿植株内木质素合成以及醌类和单宁形成, 增强冷蒿抗机械损伤能力和提高冷蒿的耐牧性。
Sesquiterpenoids, and specifically sesquiterpene lactones from Asteraceae, may play a highly significant role in human health, both as part of a balanced diet and as pharmaceutical agents, due to their potential for the treatment of cardiovascular disease and cancer. This review highlights the role of sesquiterpene lactones endogenously in the plants that produce them, and explores mechanisms by which they interact in animal and human consumers of these plants. Several mechanisms are proposed for the reduction of inflammation and tumorigenesis at potentially achievable levels in humans. Plants can be classified by their specific array of produced sesquiterpene lactones, showing high levels of translational control. Studies of folk medicines implicate sesquiterpene lactones as the active ingredient in many treatments for other ailments such as diarrhea, burns, influenza, and neurodegradation. In addition to the anti-inflammatory response, sesquiterpene lactones have been found to sensitize tumor cells to conventional drug treatments. This review explores the varied ecological roles of sesquiterpenes in the plant producer, depending upon the plant and the compound. These include allelopathy with other plants, insects, and microbes, thereby causing behavioural or developmental modification to these secondary organisms to the benefit of the sesquiterpenoid producer. Some sesquiterpenoid lactones are antimicrobial, disrupting the cell wall of fungi and invasive bacteria, whereas others protect the plant from environmental stresses that would otherwise cause oxidative damage. Many of the compounds are effective due to their bitter flavor, which has obvious implications for human consumers. The implications of sesquiterpenoid lactone qualities for future crop production are discussed.
To test the hypothesis that drought predisposes trees to insect attacks, we quantified the effects of water availability on insect attacks, tree resistance mechanisms, and mortality of mature piñon pine (Pinus edulis) and one-seed juniper (Juniperus monosperma) using an experimental drought study in New Mexico, USA. The study had four replicated treatments (40 × 40 m plot/replicate): removal of 45% of ambient annual precipitation (H2 O-); irrigation to produce 125% of ambient annual precipitation (H2 O+); a drought control (C) to quantify the impact of the drought infrastructure; and ambient precipitation (A). Piñon began dying 1 yr after drought initiation, with higher mortality in the H2 O- treatment relative to other treatments. Beetles (bark/twig) were present in 92% of dead trees. Resin duct density and area were more strongly affected by treatments and more strongly associated with piñon mortality than direct measurements of resin flow. For juniper, treatments had no effect on insect resistance or attacks, but needle browning was highest in the H2 O- treatment. Our results provide strong evidence that ≥ 1 yr of severe drought predisposes piñon to insect attacks and increases mortality, whereas 3 yr of the same drought causes partial canopy loss in juniper.© 2013 The Authors. New Phytologist © 2013 New Phytologist Trust.
Phylogenetic relationships and genomic compatibility were compared for 60 accessions of Syringa using chloroplast DNA (cpDNA) and nuclear ribosomal DNA (rDNA) markers. A total of 669 cpDNA variants, 653 of which were potentially phylogenetically informative, was detected using 22 restriction enzymes. Phylogenetic analyses reveal four strongly supported plastome groups that correspond to four genetically incompatible crossing groups. Relationships of the four plastome groups (I(II(III,IV))) correlate well with the infrageneric classification except for ser. Syringa and Pinnatifoliae. Group I, which includes subg. Ligustrina, forms a basal lineage within Syringa. Group II includes ser. Syringa and Pinnatifoliae and the two series have high compatibility and low sequence divergence. Group III consists of three well-defined species groups of ser. Pubescentes. Group IV comprises all members of ser. Villosae and has the lowest interspecific cpDNA sequence divergences. Comparison of cpDNA sequence divergence with crossability data indicates that hybrids have not been successfully generated between species with divergence greater than 0.7%. Hybrid barriers are strong among the four major plastome groups, which have sequence divergence estimates ranging from 1.096 to 1.962%. In contrast, fully fertile hybrids occur between species pairs with sequence divergence below 0.4%. Three regions of the plastome have length variants of greater than 100 bp, and these indels identify 12 different plastome types that correlate with phylogenetic trees produced from cpDNA restriction site data. Biparentally inherited nuclear rDNA and maternally inherited cpDNA length variants enable the identification of the specific parentage of several lilac hybrids.
Little is known about the mechanisms promoting or limiting the coexistence of functionally divergent species in hyperdiverse tropical tree genera. Density-dependent enemy attacks have been proposed to be a major driver for the local coexistence of chemically divergent congeneric species. At the same time, we expect local soil conditions to favor the coexistence of species sharing similar functional traits related to resource use strategies, while environmental heterogeneity would promote the diversity of these traits at both local and large spatial scales. To test how these traits mediate species coexistence, we used functional trait data for 29 species from the tree genus Protium (Burseraceae), collected in 19 plots (2 ha each) in the Peruvian Amazon. We characterized the presence-absence of 189 plant secondary metabolites (SM) for 27 of these species, and 14 functional traits associated with resource use strategies (RUT) for 16 species. Based on these data, we found that SM were significantly more dissimilar than null expectations for species co-occurring within plots, whereas RUT were significantly more similar. These results were consistent with the hypothesis that density-dependent enemy attacks contribute to the local coexistence of congeneric species displaying divergent chemical defenses, whereas local habitat conditions filter species with similar RUT. Using measurements of nine soil properties in each plot, we also found a significant turnover of RUT traits with increasing dissimilarity of soil texture and nutrient availabilities, providing support for the hypothesis that soil heterogeneity maintains functional diversity at larger spatial scales (from 500 m up to ca. 200 km) in Protium communities. Our study provides new evidence suggesting that density-dependent enemy attacks and soil heterogeneity both contribute to maintaining high species richness in diverse tropical forests.

{kind=link}
{kind=link}
{kind=link}
{kind=link}