|
|
|||||||||||||||||||||||||
|
淫羊藿类黄酮生物合成相关基因研究进展
植物学报
2024, 59 (5):
834-846.
DOI: 10.11983/CBB23133
淫羊藿(Herba Epimedii)是一种历史悠久的中草药(TCM), 药用价值高, 国内淫羊藿相关研究备受关注。黄酮醇苷类(PFGs)成分是淫羊藿的主要活性物质, 其含量决定了药用品质。了解淫羊藿类黄酮生物合成途径, 挖掘与淫羊藿黄酮醇苷类含量相关的基因, 阐明其生物合成途径的调控机制对于提高淫羊藿品质至关重要。该文系统综述了淫羊藿类黄酮生物合成相关结构基因和转录因子基因研究进展, 为揭示黄酮含量的分子调控机制, 进而为淫羊藿分子育种和黄酮醇苷的合成生物学研究奠定理论基础。
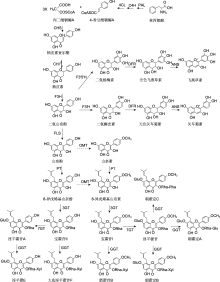
表3
淫羊藿属植物中花青素合成途径基因
正文中引用本图/表的段落
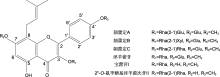
目前, 从箭叶淫羊藿、拟巫山淫羊藿和柔毛淫羊藿中克隆到1个F3′H、1个F3′5′H、2个DFR、1个ANS (Huang et al., 2012, 2015; Zeng et al., 2013a)和1个UGT编码基因(姚宇, 2023) (表3)。除UGT基因外, 均未进行功能验证。
b: 序列比对或系统发育分析; c: 异源表达和酵母双杂交检测.a及4种主要活性成分同 Isolation and molecular characterisation of flavonoid 3′-hydroxylase and flavonoid 3′,5′-hydroxylase genes from a traditional Chinese medicinal plant 4 2012 ... 目前, 从箭叶淫羊藿、拟巫山淫羊藿和柔毛淫羊藿中克隆到1个F3′H、1个F3′5′H、2个DFR、1个ANS (Huang et al.,
b: 序列比对或系统发育分析; c: 异源表达和酵母双杂交检测.a及4种主要活性成分同 Phytochemistry and ethnopharmacology of Epimedium L. species 1 2015 ... 淫羊藿(Herba Epimedii)是应用历史悠久的传统中药, 具有补肾阳、强筋骨和祛风湿等功效, 现代药理研究表明其在抗骨质疏松、抗病毒、抗肿瘤、降血糖和降血脂等方面具有显著功效(Jiang et al., MdHB1 down-regulation activates anthocyanin biosynthesis in the white-fleshed apple cultivar ‘Granny Smith’ 1 2017 ... 类黄酮在植物不同组织和器官中的种类及含量差异较大.参与其生物合成调控的因子包括转录因子、长链非编码RNA (long non-coding RNA, LncRNA)、非编码单链小RNA (microRNA, miRNA)、DNA甲基化和蛋白质泛素化等(Hu et al., Evolution and diversity of the 2-oxoglutarate-dependent dioxygenase superfamily in plants 1 2014 ... 花青素合酶(ANS)与F3H及FLS都属于2-ODDs超家族中的DOXC亚家族成员(Kawai et al., An efficient method for C8-prenylation of flavonols and flavanones 1 2012 ... 近年来, 合成生物学通过构建微生物细胞代谢途径, 已实现多个参与类黄酮生物合成的基因在微生物体内表达以及异戊烯基类黄酮的合成(Dong et al., Potential of icariin metabolites from Epimedium koreanum Nakai as antidiabetic therapeutic agents 1 2017 ... 淫羊藿(Herba Epimedii)是应用历史悠久的传统中药, 具有补肾阳、强筋骨和祛风湿等功效, 现代药理研究表明其在抗骨质疏松、抗病毒、抗肿瘤、降血糖和降血脂等方面具有显著功效(Jiang et al., PyWRKY26 and PybHLH3 cotargeted the PyMYB114 promoter to regulate anthocyanin biosynthesis and transport in red-skinned pears 1 2020 ... 类黄酮在植物不同组织和器官中的种类及含量差异较大.参与其生物合成调控的因子包括转录因子、长链非编码RNA (long non-coding RNA, LncRNA)、非编码单链小RNA (microRNA, miRNA)、DNA甲基化和蛋白质泛素化等(Hu et al., Advances of synthetic biology of flavonoids 1 2022 ... 从苯丙氨酸到柚皮素(二氢黄酮)是所有已知类黄酮化合物生物合成的共同上游途径(林春草等, 黄酮类化合物合成生物学研究进展 1 2022 ... 从苯丙氨酸到柚皮素(二氢黄酮)是所有已知类黄酮化合物生物合成的共同上游途径(林春草等, MYB transcription factors as regulators of phenylpropanoid metabolism in plants 1 2015 ... 类黄酮在植物不同组织和器官中的种类及含量差异较大.参与其生物合成调控的因子包括转录因子、长链非编码RNA (long non-coding RNA, LncRNA)、非编码单链小RNA (microRNA, miRNA)、DNA甲基化和蛋白质泛素化等(Hu et al., Identification and characterization of long non-coding RNAs regulating flavonoid biosynthesis in Ginkgo biloba leaves 1 2020 ... 类黄酮在植物不同组织和器官中的种类及含量差异较大.参与其生物合成调控的因子包括转录因子、长链非编码RNA (long non-coding RNA, LncRNA)、非编码单链小RNA (microRNA, miRNA)、DNA甲基化和蛋白质泛素化等(Hu et al., The flavonoid biosynthesis network in plants 1 2021a ... 从苯丙氨酸到柚皮素(二氢黄酮)是所有已知类黄酮化合物生物合成的共同上游途径(林春草等, 7 2021b ... 目前, 在箭叶淫羊藿、柔毛淫羊藿、巫山淫羊藿(E. wu- shanense)和拟巫山淫羊藿(E. pseudowushanense)中发现了5个PAL、2个C4H、6个4CL、6个CHS、6个CHI和2个F3H编码基因(Zeng et al.,
本文的其它图/表
|
 首页
首页