高温胁迫对植物光合作用的影响研究进展
孙永江, 王琪, 邵琪雯, 辛智鸣, 肖辉杰, 程瑾
植物学报
2023, 58 ( 3):
486-498.
DOI: 10.11983/CBB22079
随着人为活动产生的大气CO2浓度的增加, 全球气候持续变暖。过去5年是自有温度记录以来最热的5年。高温胁迫已经成为影响植物生长发育的主要逆境因子之一。光合作用是地球生命活动的基础, 对环境波动高度敏感。解析植物在高温环境下光合作用的响应特性, 可为探索植物抵御高温的生理生态机制、培育抗高温新品种以及采取合理措施适应未来极端气候提供科学依据。该文论述了高温胁迫对植物光合电子传递及碳固定过程的影响, 从光质和光强角度综合分析了光照对高温胁迫下光合作用的影响; 从植物自身及外源缓解物质等方面阐述了植物增强抗高温胁迫的途径和机制。同时, 对植物光合作用响应高温胁迫的研究方向及多组学联合分析在揭示植物抵御高温胁迫机制中的应用进行了展望。
| 植物种名 | 外源物质 | 处理温度/时间 | 浓度/方式 | 参考文献 | | 烟草(Nicotiana tabacum) | 氯化钙 | 40°C/24小时 | 20 mmol?L-1/叶面喷施 | Tan et al., 2011 | | 水稻(Oryza sativa) | 表油菜素内酯 | 40°C/7天 | 0.1 μmol?L-1/叶面喷施 | Thussagunpanit et al., 2015 | | 马铃薯(Solanum tuberosum) | 亚磷酸钾 | 37°C/24小时 | 1%/土壤施入 | Xi et al., 2020 | | 番茄(S. lycopersicum) | 褪黑素 | 42°C/24小时 | 100 mmol?L-1/叶面喷施 | Jahan et al., 2021 | | 小麦(Triticum aestivum) | 海藻糖 | 40°C/24小时 | 1.5 mmol?L-1/水培施入 | Luo et al., 2021 | | 水稻(O. sativa) | 亚精胺 | 37.5°C/22天 | 1 mmol?L-1/叶面喷施 | Tang et al., 2018 | | 高羊茅(Lolium arundinaceum) | 柠檬酸 | 35°C/15天 | 20 mmol?L-1/叶面喷施 | Hu et al., 2016 | | 番茄(S. lycopersicum) | 抗坏血酸 | 40°C/8小时 | 0.5 mmol?L-1/叶面喷施 | Alayafi, 2020 |
View table in article
表2
高温胁迫下外源物质对植物光合作用的缓解
正文中引用本图/表的段落
研究发现, CEF的重要生理功能是参与非光化学淬灭(NPQ)的诱导、PSI复合体的氧化还原调控以及ATP/NADPH比例的调节(黄伟等, 2012; Suorsa et al., 2016)。NPQ是植物进行热耗散的主要机制(Müller et al., 2001; Nishiyama et al., 2001), 依赖于跨膜质子梯度(proton gradient, ΔpH)的高能态荧光淬灭(qE)是NPQ的主要组成部分(Ruban, 2016)。而逆境下CEF是叶绿体中ΔpH形成的主要来源(Aihara et al., 2016)。高温胁迫下, 植物激活CEF, 通过促进ΔpH使较高的qE热耗散(Sun et al., 2017) (图2)。此外, 有证据表明H2O2可能作为环境胁迫和CEF诱导的连接因素, 并在激活CEF过程中起作用(Strand et al., 2015)。Ca2+在此过程中也发挥重要作用(Lascano et al., 2003)。综合前人的研究结果, 推测高温胁迫主要通过引起过剩光能积累导致ROS产生, ROS进而作为信号分子, 通过Ca2+信号级联反应诱导CEF活性。
(A) 光合电子和质子的定向转移依赖于类囊体膜中的关键蛋白复合体; 适宜环境温度下, PSII裂解H2O中的电子主要通过线性电子传递(LEF)至光系统I, 在Fd和FNR的作用下, 将NADP+还原成NADPH, 形成固定CO2所需的还原力; (B) 高温胁迫下, LEF被抑制, PQ库被还原的同时, 依赖NDH及质子梯度调节蛋白/质子梯度调节类似蛋白(PGR5/PGRL1)的环式电子传递(CEF)被激活; LEF的下调可以通过降低传递至光系统I处的电子促进P700的氧化, 而CEF的上调可以通过增加跨膜质子梯度来产生更多的ATP, 也可以通过激发非光化学淬灭来保护光合机构; 因此, 高温胁迫下, LEF和CEF协同作用调节PQ库及P700的氧化还原状态及热耗散, 起到光保护作用。P680和P700分别表示PSII和PSI反应中心叶绿素在可见光谱中的波长吸收峰值。绿色箭头表示LEF转移, 红线表示CEF转移。Fd: 铁氧还蛋白; FNR: 黄素蛋白铁氧还蛋白-NADP还原酶; NADP+: 氧化型烟酰胺腺嘌呤二核苷磷酸; NDH: NA(P)H脱氢酶复合体; PQ: 质体醌; PQH2: 质体醌醇
合理施用外源物质是调节植物生长发育并应对不利环境的重要栽培措施, 能达到调节植物高温胁迫抵御能力的效果。表2列举了具有缓解高温胁迫的外源物质及施用条件。其中, Ca2+作为在植物生长发育过程中的必需元素, 是植物体内重要的第二信使, 参与调节植物对各种非生物胁迫的响应(Hashimoto and Kudla, 2011)。外源Ca2+可以减少活性氧的积累, 通过改善叶片气孔导度及提高放氧复合体(OEC)的热稳定性, 改善高温胁迫下植物的光合作用(Tan et al., 2011)。RNA测序发现类钙调素蛋白(CML)基因可能对高温胁迫产生积极响应, 调节植物抵御高温胁迫(Wang et al., 2019)。
亚磷酸盐(Phi)作为环境友好型外源物质而备受关注(Mayton et al., 2008)。外源施加亚磷酸钾可增强抗氧化酶的活性, 减少H2O2积累, 上调光合基因表达水平, 并诱导葡聚糖酶和几丁质酶的积累, 进而阻止UV-B对光合作用的破坏(Oyarburo et al., 2015)。Phi还诱导应激蛋白, 促进渗透物质的积累, 缓解DNA损伤, 提高植物抵御高温胁迫的能力(Xi et al., 2020)。此外, 还发现外源褪黑素、海藻糖、亚精胺、柠檬酸及抗坏血酸可缓解高温胁迫对植物的影响, 施用浓度和方式见表2。
Rubisco: 1,5-二磷酸核酮糖羧化酶/加氧酶; PEPC: 磷酸烯醇式丙酮酸羧化酶; SBPase: 景天庚酮糖1,6-二磷酸酯酶; TK: 转酮醇酶; PRK: 磷酸核酮糖激酶 ... Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants 1 2013 ... 叶绿体类囊体膜的结构和功能变化影响光合作用过程, 环境温度的变化导致类囊体膜的流动性增强, 类囊体膜组分发生相变, 从而影响膜稳定性.严重高温胁迫会破坏叶绿体被膜的完整性(Gounaris et al., 1984).此外, 高温胁迫易造成叶片总叶绿素含量降低(Hasanuzzaman et al., 2013).叶绿素作为叶绿体类囊体膜上的主要光合色素, 起到捕获并驱动光合电子传递的作用.在正常条件下, 叶绿素的合成与降解处于平衡状态.当植物处于高温环境中, 一方面参与吡咯合成途径的第1个酶(5-氨基戊酮酸脱水酶)活性被抑制(Mohanty et al., 2006); 另一方面加剧了叶绿素酶和叶绿素降解过氧化物酶活性降低(Wang et al., 2018), 导致叶绿素含量降低, 引起叶片衰老.由于叶绿素捕获过多光能将威胁到植物的光合作用, 因此叶绿素的降解在一定程度上也是植物的一种保护反应. ... Calcium decoding mechanisms in plants 1 2011 ... 合理施用外源物质是调节植物生长发育并应对不利环境的重要栽培措施, 能达到调节植物高温胁迫抵御能力的效果.表2列举了具有缓解高温胁迫的外源物质及施用条件.其中, Ca2+作为在植物生长发育过程中的必需元素, 是植物体内重要的第二信使, 参与调节植物对各种非生物胁迫的响应(Hashimoto and Kudla, 2011).外源Ca2+可以减少活性氧的积累, 通过改善叶片气孔导度及提高放氧复合体(OEC)的热稳定性, 改善高温胁迫下植物的光合作用(Tan et al., 2011).RNA测序发现类钙调素蛋白(CML)基因可能对高温胁迫产生积极响应, 调节植物抵御高温胁迫(Wang et al., 2019). ... Functioning of photosystems I and II in pea leaves exposed to heat stress in the presence or absence of light: analysis using in vivo fluorescence, absorbance, oxygen and photoa- coustic measurements 1 1991 ... 光照不足会导致同化力减弱和光合作用的关键酶未能充分活化进而限制光合作用; 而光强过高会造成植物吸收超过其自身能够利用的光能, 导致过剩激发能的累积, 不可避免地产生ROS, 引发严重的光抑制(Takahashi and Murata, 2008).为了应对时刻变化的光照环境, 植物通过调节光合电子传递产物ATP以及NADPH来适应碳同化的消耗(Foyer et al., 2012).此外, 植物还通过编码2个光系统反应中心亚基的基因快速响应强光(Murchie et al., 2005).高温黑暗对植物PSII活性的伤害已被广泛研究(Li et al., 2009; Xu et al., 2014; Essemine et al., 2016).有趣的是, 研究发现光对高温胁迫下植物光系统具有保护作用(Havaux et al., 1991; Marutani et al., 2012; Buchner et al., 2015).光强直接影响D1蛋白的周转(Tyystj?rvi and Aro, 1996).在一定光强范围内, 随着光强的增加, D1蛋白的周转速度加快, 在中等光强下达到最大值(Russell et al., 1995).Rubisco大、小亚基的含量在黑暗下降低, 而在光下则增高(Demire- vska-Kepova et al., 2005).RCA活性也随着光强的增加而增强(姜振升等, 2010), 从而可能对RCA基因表达进行转录后调控(DeRidder and Salvucci, 2007).研究发现CEF途径受光照影响(Teicher and Scheller, 1998).弱光下CEF的增加对水稻抵御夏季高温逆境具有重要作用(Yamori et al., 2015).这种机制对于探究通过提高植物光合作用抵御高温胁迫具有重要意义. ... Function of the two cytochrome components in chloroplasts: a working hypothesis 1 1960 ... 正常条件下, 植物光合作用存在至少2种电子传递途径, 即由PSII、PQ库、细胞色素(Cyt)b6/f复合体和PSI推动的线性电子传递(linear electron flow, LEF) (Hill and Bendall, 1960)与仅由PSI推动的环式电子传递(cyclic electron flow, CEF) (Munekage et al., 2004; 刘玉凤等, 2019).逆境胁迫下LEF的降低能够避免过量电子从PSII传递至PSI, 从而缓解ROS在PSI的积累, 保护PSI免受高温损伤(Tikkanen et al., 2014; Jiang et al., 2021).在光合电子传递中, PQ库在PSII与Cyt b6/f复合体之间起功能连接作用, 其氧化还原状态可有效调节光合反应, 包括调节状态转换、叶绿素生物合成及光系统蛋白质合成速率(Allen, 1995; Maxwell et al., 1995).由于高温胁迫对OEC造成损伤, 导致叶片氧化态PQ库比例增加(Pshybytko et al., 2008), 而氧化态PQ库可有效淬灭PSI天线中处于激发态的叶绿素分子(Joly and Carpentier, 2007), 发挥保护PSI的作用. ... Exogenous application of citric acid ameliorates the adverse effect of heat stress in tall fescue (Lolium arundinaceum) 1 2016 ... Alleviation of exogenous substances to photosynthesis under high temperature stress
油菜素内酯(brassinosteroids, BR)影响植物对各种生物和非生物胁迫的耐受性并调控植物的生长发育.BR在HSP诱导中发挥重要作用(Sadura et al., 2020), 并增强抗氧化酶活性(Cao and Zhao, 2008).油菜素类固醇反应的关键调节因子(BZR1)参与BR介导的植物发育过程, 通过调控活性氧信号代谢通路响应高温胁迫(Yin et al., 2018).研究发现, 高温胁迫下外施BR通过提高植物叶片羧化效率和增强抗氧化系统功能进而缓解光抑制(Ogweno et al., 2008), 并促进气孔导度增大, 维持叶片CO2固定速率(Thus sagunpanit et al., 2015). ... Damage to photosystem II due to heat stress without light-driven electron flow: involvement of enhanced introduction of reducing power into thylakoid membranes 1 2012 ... 光照不足会导致同化力减弱和光合作用的关键酶未能充分活化进而限制光合作用; 而光强过高会造成植物吸收超过其自身能够利用的光能, 导致过剩激发能的累积, 不可避免地产生ROS, 引发严重的光抑制(Takahashi and Murata, 2008).为了应对时刻变化的光照环境, 植物通过调节光合电子传递产物ATP以及NADPH来适应碳同化的消耗(Foyer et al., 2012).此外, 植物还通过编码2个光系统反应中心亚基的基因快速响应强光(Murchie et al., 2005).高温黑暗对植物PSII活性的伤害已被广泛研究(Li et al., 2009; Xu et al., 2014; Essemine et al., 2016).有趣的是, 研究发现光对高温胁迫下植物光系统具有保护作用(Havaux et al., 1991; Marutani et al., 2012; Buchner et al., 2015).光强直接影响D1蛋白的周转(Tyystj?rvi and Aro, 1996).在一定光强范围内, 随着光强的增加, D1蛋白的周转速度加快, 在中等光强下达到最大值(Russell et al., 1995).Rubisco大、小亚基的含量在黑暗下降低, 而在光下则增高(Demire- vska-Kepova et al., 2005).RCA活性也随着光强的增加而增强(姜振升等, 2010), 从而可能对RCA基因表达进行转录后调控(DeRidder and Salvucci, 2007).研究发现CEF途径受光照影响(Teicher and Scheller, 1998).弱光下CEF的增加对水稻抵御夏季高温逆境具有重要作用(Yamori et al., 2015).这种机制对于探究通过提高植物光合作用抵御高温胁迫具有重要意义. ... Analysis of high temperature stress on the dynamics of antenna size and reducing side heterogeneity of photosystem II in wheat (Triticum aestivum) leaves 1 2011 ... 植物光系统由光系统I (photosystem I, PSI)和光系统II (PSII)构成.PSII是光合机构中对高温胁迫最敏感的部分之一(Berry and Bjorkman, 1980), 是光抑制的主要发生部位.高温胁迫下, 由于类囊体膜的流动性增强, PSII捕光复合体很容易从类囊体膜上脱落, 从而破坏PSII的完整性, 影响光合电子传递(Mathur et al., 2011).高温胁迫对PSII放氧复合体(oxygen evolving complex, OEC) (Tyystj?rvi, 2012)、反应中心及电子受体侧(Wise et al., 2004)均产生抑制作用, 其抑制程度主要取决于光破坏和光修复的速率.如果PSII的破坏速率超过修复速率, 将导致光能利用效率降低, 造成光抑制(Takahashi and Badger, 2011).高温胁迫下, 当光合机构吸收的过剩激发能不能被及时耗散时, 容易引发活性氧(reactive oxygen species, ROS)累积, 一方面直接破坏PSII反应中心蛋白; 另一方面通过对D1蛋白的从头合成阻碍PSII修复过程, 从而加剧PSII光抑制(Takahashi and Badger, 2011) (图1). ... Redox regulation of light-harvesting complex II and cab mRNA abundance in Dunaliella salina 1 1995 ... 正常条件下, 植物光合作用存在至少2种电子传递途径, 即由PSII、PQ库、细胞色素(Cyt)b6/f复合体和PSI推动的线性电子传递(linear electron flow, LEF) (Hill and Bendall, 1960)与仅由PSI推动的环式电子传递(cyclic electron flow, CEF) (Munekage et al., 2004; 刘玉凤等, 2019).逆境胁迫下LEF的降低能够避免过量电子从PSII传递至PSI, 从而缓解ROS在PSI的积累, 保护PSI免受高温损伤(Tikkanen et al., 2014; Jiang et al., 2021).在光合电子传递中, PQ库在PSII与Cyt b6/f复合体之间起功能连接作用, 其氧化还原状态可有效调节光合反应, 包括调节状态转换、叶绿素生物合成及光系统蛋白质合成速率(Allen, 1995; Maxwell et al., 1995).由于高温胁迫对OEC造成损伤, 导致叶片氧化态PQ库比例增加(Pshybytko et al., 2008), 而氧化态PQ库可有效淬灭PSI天线中处于激发态的叶绿素分子(Joly and Carpentier, 2007), 发挥保护PSI的作用. ... Potato late blight in tubers—the role of foliar phosphonate applications in suppressing pre-harvest tuber infections 1 2008 ... 亚磷酸盐(Phi)作为环境友好型外源物质而备受关注(Mayton et al., 2008).外源施加亚磷酸钾可增强抗氧化酶的活性, 减少H2O2积累, 上调光合基因表达水平, 并诱导葡聚糖酶和几丁质酶的积累, 进而阻止UV-B对光合作用的破坏(Oyarburo et al., 2015).Phi还诱导应激蛋白, 促进渗透物质的积累, 缓解DNA损伤, 提高植物抵御高温胁迫的能力(Xi et al., 2020).此外, 还发现外源褪黑素、海藻糖、亚精胺、柠檬酸及抗坏血酸可缓解高温胁迫对植物的影响, 施用浓度和方式见表2. ... How do plants feel the heat? 1 2012 ... 植物对高温等非生物胁迫响应的分子机制包括感知、信号转导、转录、转录处理、翻译和翻译后蛋白质修饰等多个过程(Zhang et al., 2022).植物感知高温及信号转导对光合作用的稳定至关重要.环境温度升高通过增加质膜的流动性激活Ca2+通道, 促进Ca2+内流至细胞质, 产生胞内信号(Sajid et al., 2018), 并继续通过Ca2+、ROS和不同类型的蛋白激酶等第二信使传递并放大, 引起转录因子的激活和植物激素的级联控制反应(Mittler et al., 2012), 进而激活包括热激蛋白(HSPs)等在内的相对保守的调控网络(Richter et al., 2010), 稳定高温胁迫下叶绿体的发育和PSII的功能(Zhong et al., 2013).而热激因子(heat shock factor, HSF)作为ROS的感受器(Charng et al., 2007), 在调控HSPs表达及参与受损蛋白修复的过程中起关键作用(Qu et al., 2013).位于叶绿体的HSP21被GUN5 (GENOMES UNCOUPLED 5)依赖的逆向信号通路激活, 并通过直接结合D1和D2蛋白等核心亚基稳定高温胁迫下PSII的功能(Chen et al., 2017).此外, 在热适应提高植物高温胁迫耐受性过程中, 植物对热胁迫的记忆依赖于HSPs诱导(Wu et al., 2013)及叶绿体中Ca2+浓度的升高(Pollastri et al., 2021). ... Dynamic mechanical and thermal properties of MAPE treated jute/ HDPE composites 1 2006 ... 叶绿体类囊体膜的结构和功能变化影响光合作用过程, 环境温度的变化导致类囊体膜的流动性增强, 类囊体膜组分发生相变, 从而影响膜稳定性.严重高温胁迫会破坏叶绿体被膜的完整性(Gounaris et al., 1984).此外, 高温胁迫易造成叶片总叶绿素含量降低(Hasanuzzaman et al., 2013).叶绿素作为叶绿体类囊体膜上的主要光合色素, 起到捕获并驱动光合电子传递的作用.在正常条件下, 叶绿素的合成与降解处于平衡状态.当植物处于高温环境中, 一方面参与吡咯合成途径的第1个酶(5-氨基戊酮酸脱水酶)活性被抑制(Mohanty et al., 2006); 另一方面加剧了叶绿素酶和叶绿素降解过氧化物酶活性降低(Wang et al., 2018), 导致叶绿素含量降低, 引起叶片衰老.由于叶绿素捕获过多光能将威胁到植物的光合作用, 因此叶绿素的降解在一定程度上也是植物的一种保护反应. ... Non-photochemical quenching. A response to excess light energy 1 2001 ... 研究发现, CEF的重要生理功能是参与非光化学淬灭(NPQ)的诱导、PSI复合体的氧化还原调控以及ATP/NADPH比例的调节(黄伟等, 2012; Suorsa et al., 2016).NPQ是植物进行热耗散的主要机制(Müller et al., 2001; Nishiyama et al., 2001), 依赖于跨膜质子梯度(proton gradient, ΔpH)的高能态荧光淬灭(qE)是NPQ的主要组成部分(Ruban, 2016).而逆境下CEF是叶绿体中ΔpH形成的主要来源(Aihara et al., 2016).高温胁迫下, 植物激活CEF, 通过促进ΔpH使较高的qE热耗散(Sun et al., 2017) (图2).此外, 有证据表明H2O2可能作为环境胁迫和CEF诱导的连接因素, 并在激活CEF过程中起作用(Strand et al., 2015).Ca2+在此过程中也发挥重要作用(Lascano et al., 2003).综合前人的研究结果, 推测高温胁迫主要通过引起过剩光能积累导致ROS产生, ROS进而作为信号分子, 通过Ca2+信号级联反应诱导CEF活性. ... Cyclic electron flow around photosystem I is essential for photosynthesis 1 2004 ... 正常条件下, 植物光合作用存在至少2种电子传递途径, 即由PSII、PQ库、细胞色素(Cyt)b6/f复合体和PSI推动的线性电子传递(linear electron flow, LEF) (Hill and Bendall, 1960)与仅由PSI推动的环式电子传递(cyclic electron flow, CEF) (Munekage et al., 2004; 刘玉凤等, 2019).逆境胁迫下LEF的降低能够避免过量电子从PSII传递至PSI, 从而缓解ROS在PSI的积累, 保护PSI免受高温损伤(Tikkanen et al., 2014; Jiang et al., 2021).在光合电子传递中, PQ库在PSII与Cyt b6/f复合体之间起功能连接作用, 其氧化还原状态可有效调节光合反应, 包括调节状态转换、叶绿素生物合成及光系统蛋白质合成速率(Allen, 1995; Maxwell et al., 1995).由于高温胁迫对OEC造成损伤, 导致叶片氧化态PQ库比例增加(Pshybytko et al., 2008), 而氧化态PQ库可有效淬灭PSI天线中处于激发态的叶绿素分子(Joly and Carpentier, 2007), 发挥保护PSI的作用. ... Acclimation of photosynthesis to high irradiance in rice: gene expression and interactions with leaf development 1 2005 ... 光照不足会导致同化力减弱和光合作用的关键酶未能充分活化进而限制光合作用; 而光强过高会造成植物吸收超过其自身能够利用的光能, 导致过剩激发能的累积, 不可避免地产生ROS, 引发严重的光抑制(Takahashi and Murata, 2008).为了应对时刻变化的光照环境, 植物通过调节光合电子传递产物ATP以及NADPH来适应碳同化的消耗(Foyer et al., 2012).此外, 植物还通过编码2个光系统反应中心亚基的基因快速响应强光(Murchie et al., 2005).高温黑暗对植物PSII活性的伤害已被广泛研究(Li et al., 2009; Xu et al., 2014; Essemine et al., 2016).有趣的是, 研究发现光对高温胁迫下植物光系统具有保护作用(Havaux et al., 1991; Marutani et al., 2012; Buchner et al., 2015).光强直接影响D1蛋白的周转(Tyystj?rvi and Aro, 1996).在一定光强范围内, 随着光强的增加, D1蛋白的周转速度加快, 在中等光强下达到最大值(Russell et al., 1995).Rubisco大、小亚基的含量在黑暗下降低, 而在光下则增高(Demire- vska-Kepova et al., 2005).RCA活性也随着光强的增加而增强(姜振升等, 2010), 从而可能对RCA基因表达进行转录后调控(DeRidder and Salvucci, 2007).研究发现CEF途径受光照影响(Teicher and Scheller, 1998).弱光下CEF的增加对水稻抵御夏季高温逆境具有重要作用(Yamori et al., 2015).这种机制对于探究通过提高植物光合作用抵御高温胁迫具有重要意义. ... A new paradigm for the action of reactive oxygen species in the photoinhibition of photosystem II. 1 2006 ... 在高温胁迫下, 植物采取的光保护机制包括2方面: 避免暴露在强光环境及合理应对过剩光能(Takahashi and Badger, 2011).从整株尺度上, 植物通过树冠内相互遮蔽或调节叶片角度减少受辐射强度, 以及通过蒸腾作用等途径降低叶温(Tserej and Feeley, 2021).由于气孔导度的瞬态增加, 导致蒸腾作用增大, 而蒸腾作用是植物主要降温机制; 此外, 通过调节小气候, 引起同一林分内不同物种甚至同一树冠内不同叶片之间的叶温出现明显差异, 维持大部分叶片的叶温处于光合作用的适宜温度范围内, 避免受极端高温的损害(Fauset et al., 2018).在叶片尺度上避免高温伤害的策略有叶片运动(Kao and Forseth, 1992)和叶绿体运动(Wada et al., 2003), 从而避免叶片和叶绿体受到过强光照, 保护光合机构免受高温伤害; 耐热策略则主要有活性氧清除(Nishiyama et al., 2006)、将捕光复合体吸收的过剩光能以热能的形式耗散(Sherstn- eva et al., 2022)、光合电子传递调节(Jiang et al., 2021)及光呼吸(光下植物吸收O2并释放CO2) (Osei- Bonsu et al., 2021)等. ... Oxidative stress inhibits the repair of photodamage to the photosynthetic machinery 1 2001 ... 研究发现, CEF的重要生理功能是参与非光化学淬灭(NPQ)的诱导、PSI复合体的氧化还原调控以及ATP/NADPH比例的调节(黄伟等, 2012; Suorsa et al., 2016).NPQ是植物进行热耗散的主要机制(Müller et al., 2001; Nishiyama et al., 2001), 依赖于跨膜质子梯度(proton gradient, ΔpH)的高能态荧光淬灭(qE)是NPQ的主要组成部分(Ruban, 2016).而逆境下CEF是叶绿体中ΔpH形成的主要来源(Aihara et al., 2016).高温胁迫下, 植物激活CEF, 通过促进ΔpH使较高的qE热耗散(Sun et al., 2017) (图2).此外, 有证据表明H2O2可能作为环境胁迫和CEF诱导的连接因素, 并在激活CEF过程中起作用(Strand et al., 2015).Ca2+在此过程中也发挥重要作用(Lascano et al., 2003).综合前人的研究结果, 推测高温胁迫主要通过引起过剩光能积累导致ROS产生, ROS进而作为信号分子, 通过Ca2+信号级联反应诱导CEF活性. ... Brassinosteroids alleviate heat-induced inhibition of photosynthesis by increasing carboxylation efficiency and enhancing antioxidant systems in Lycopersicon esculentum 1 2008 ... 油菜素内酯(brassinosteroids, BR)影响植物对各种生物和非生物胁迫的耐受性并调控植物的生长发育.BR在HSP诱导中发挥重要作用(Sadura et al., 2020), 并增强抗氧化酶活性(Cao and Zhao, 2008).油菜素类固醇反应的关键调节因子(BZR1)参与BR介导的植物发育过程, 通过调控活性氧信号代谢通路响应高温胁迫(Yin et al., 2018).研究发现, 高温胁迫下外施BR通过提高植物叶片羧化效率和增强抗氧化系统功能进而缓解光抑制(Ogweno et al., 2008), 并促进气孔导度增大, 维持叶片CO2固定速率(Thus sagunpanit et al., 2015). ... The roles of photorespiration and alternative electron acceptors in the responses of photosynthesis to elevated temperatures in cowpea 1 2021 ... 在高温胁迫下, 植物采取的光保护机制包括2方面: 避免暴露在强光环境及合理应对过剩光能(Takahashi and Badger, 2011).从整株尺度上, 植物通过树冠内相互遮蔽或调节叶片角度减少受辐射强度, 以及通过蒸腾作用等途径降低叶温(Tserej and Feeley, 2021).由于气孔导度的瞬态增加, 导致蒸腾作用增大, 而蒸腾作用是植物主要降温机制; 此外, 通过调节小气候, 引起同一林分内不同物种甚至同一树冠内不同叶片之间的叶温出现明显差异, 维持大部分叶片的叶温处于光合作用的适宜温度范围内, 避免受极端高温的损害(Fauset et al., 2018).在叶片尺度上避免高温伤害的策略有叶片运动(Kao and Forseth, 1992)和叶绿体运动(Wada et al., 2003), 从而避免叶片和叶绿体受到过强光照, 保护光合机构免受高温伤害; 耐热策略则主要有活性氧清除(Nishiyama et al., 2006)、将捕光复合体吸收的过剩光能以热能的形式耗散(Sherstn- eva et al., 2022)、光合电子传递调节(Jiang et al., 2021)及光呼吸(光下植物吸收O2并释放CO2) (Osei- Bonsu et al., 2021)等. ... Potassium phos- phite increases tolerance to UV-B in potato 1 2015 ... 亚磷酸盐(Phi)作为环境友好型外源物质而备受关注(Mayton et al., 2008).外源施加亚磷酸钾可增强抗氧化酶的活性, 减少H2O2积累, 上调光合基因表达水平, 并诱导葡聚糖酶和几丁质酶的积累, 进而阻止UV-B对光合作用的破坏(Oyarburo et al., 2015).Phi还诱导应激蛋白, 促进渗透物质的积累, 缓解DNA损伤, 提高植物抵御高温胁迫的能力(Xi et al., 2020).此外, 还发现外源褪黑素、海藻糖、亚精胺、柠檬酸及抗坏血酸可缓解高温胁迫对植物的影响, 施用浓度和方式见表2. ... Increasing trends in regional heatwaves 1 2020 ... 光合作用是地球上植物生命活动的能量和物质基础.作为自然界中最重要的生物物理及生物化学反应之一, 光合作用对环境波动高度敏感, 通过诊断光合机构运转状况, 可以判断植物对环境的耐受能力(许大全, 2002).近年来, 由于人为活动导致大气中CO2浓度持续增加, 全球温室效应加剧, 预计气候变暖造成的高温胁迫将日益频繁和强烈(Perkins-Kirkpatrick and Lewis, 2020), 给现代农林业生产体系带来严峻的挑战(Tarvainen et al., 2022).2011-2020年的10年间, 全球地表温度比1850-1900年间高1.09°C, 过去5年也是自1850年有温度记录以来最热的5年(IPCC, 2021).尽管2020年新型冠状病毒(COVID-19)流行期间相关人为活动减少导致碳排放量有所下降(Le Quéré et al., 2020), 但这样的改善是短暂的, 全球气温将持续升高(Tollefson, 2021). ... Chloroplast calcium signaling regulates thermomemory 1 2021 ... 植物对高温等非生物胁迫响应的分子机制包括感知、信号转导、转录、转录处理、翻译和翻译后蛋白质修饰等多个过程(Zhang et al., 2022).植物感知高温及信号转导对光合作用的稳定至关重要.环境温度升高通过增加质膜的流动性激活Ca2+通道, 促进Ca2+内流至细胞质, 产生胞内信号(Sajid et al., 2018), 并继续通过Ca2+、ROS和不同类型的蛋白激酶等第二信使传递并放大, 引起转录因子的激活和植物激素的级联控制反应(Mittler et al., 2012), 进而激活包括热激蛋白(HSPs)等在内的相对保守的调控网络(Richter et al., 2010), 稳定高温胁迫下叶绿体的发育和PSII的功能(Zhong et al., 2013).而热激因子(heat shock factor, HSF)作为ROS的感受器(Charng et al., 2007), 在调控HSPs表达及参与受损蛋白修复的过程中起关键作用(Qu et al., 2013).位于叶绿体的HSP21被GUN5 (GENOMES UNCOUPLED 5)依赖的逆向信号通路激活, 并通过直接结合D1和D2蛋白等核心亚基稳定高温胁迫下PSII的功能(Chen et al., 2017).此外, 在热适应提高植物高温胁迫耐受性过程中, 植物对热胁迫的记忆依赖于HSPs诱导(Wu et al., 2013)及叶绿体中Ca2+浓度的升高(Pollastri et al., 2021). ... Function of plastoquinone in heat stress reactions of plants 1 2008 ... 正常条件下, 植物光合作用存在至少2种电子传递途径, 即由PSII、PQ库、细胞色素(Cyt)b6/f复合体和PSI推动的线性电子传递(linear electron flow, LEF) (Hill and Bendall, 1960)与仅由PSI推动的环式电子传递(cyclic electron flow, CEF) (Munekage et al., 2004; 刘玉凤等, 2019).逆境胁迫下LEF的降低能够避免过量电子从PSII传递至PSI, 从而缓解ROS在PSI的积累, 保护PSI免受高温损伤(Tikkanen et al., 2014; Jiang et al., 2021).在光合电子传递中, PQ库在PSII与Cyt b6/f复合体之间起功能连接作用, 其氧化还原状态可有效调节光合反应, 包括调节状态转换、叶绿素生物合成及光系统蛋白质合成速率(Allen, 1995; Maxwell et al., 1995).由于高温胁迫对OEC造成损伤, 导致叶片氧化态PQ库比例增加(Pshybytko et al., 2008), 而氧化态PQ库可有效淬灭PSI天线中处于激发态的叶绿素分子(Joly and Carpentier, 2007), 发挥保护PSI的作用. ... Protein modification in plants in response to abiotic stress 1 2019 ... 翻译后修饰是植物快速调节底物蛋白性质和活性的重要机制(Qazi et al., 2019), 也参与逆境胁迫下叶绿体蛋白的调控(Grabsztunowicz et al., 2017).研究发现, SUMO化作为重要的翻译后修饰机制, 能够参与调节植物细胞的热胁迫反应(Rytz et al., 2018).叶绿体是植物细胞中对高温最敏感的细胞器, 为应对高温胁迫, 叶绿体内发生大规模的蛋白质组重塑.叶绿体中的大量蛋白由核基因组编码, 高温导致叶绿体蛋白出现错误定位.SUMO化修饰可以通过蛋白磷酸化正向调节前体蛋白向叶绿体转运的效率, 有助于维持其在叶绿体的定位(Zheng et al., 2022). ... Molecular mechanisms of the plant heat stress response 1 2013 ... 植物对高温等非生物胁迫响应的分子机制包括感知、信号转导、转录、转录处理、翻译和翻译后蛋白质修饰等多个过程(Zhang et al., 2022).植物感知高温及信号转导对光合作用的稳定至关重要.环境温度升高通过增加质膜的流动性激活Ca2+通道, 促进Ca2+内流至细胞质, 产生胞内信号(Sajid et al., 2018), 并继续通过Ca2+、ROS和不同类型的蛋白激酶等第二信使传递并放大, 引起转录因子的激活和植物激素的级联控制反应(Mittler et al., 2012), 进而激活包括热激蛋白(HSPs)等在内的相对保守的调控网络(Richter et al., 2010), 稳定高温胁迫下叶绿体的发育和PSII的功能(Zhong et al., 2013).而热激因子(heat shock factor, HSF)作为ROS的感受器(Charng et al., 2007), 在调控HSPs表达及参与受损蛋白修复的过程中起关键作用(Qu et al., 2013).位于叶绿体的HSP21被GUN5 (GENOMES UNCOUPLED 5)依赖的逆向信号通路激活, 并通过直接结合D1和D2蛋白等核心亚基稳定高温胁迫下PSII的功能(Chen et al., 2017).此外, 在热适应提高植物高温胁迫耐受性过程中, 植物对热胁迫的记忆依赖于HSPs诱导(Wu et al., 2013)及叶绿体中Ca2+浓度的升高(Pollastri et al., 2021). ... Molecular and genetic control of plant thermomorphogenesis 1 2016 ... 光质主要通过植物感光器对高温胁迫响应起作用.研究表明, 植物体内光、温信号存在分子联系, 其中光敏色素B (PhyB)不仅起光受体的作用, 还具有温度受体的作用.远红光(FR)和适度高温使PhyB失活(Jung et al., 2016; Legris et al., 2017).而拟南芥PhyB突变体的耐热性增强(Song et al., 2017), 表明PhyB在植物适应高温过程中起重要作用.此外, 光敏色素相互作用因子(PIFs)等转录因子在光信号和热形态建成中均发挥作用(Lau et al., 2018).与自然光下生长的拟南芥相比, 在低红光/远红光比率(L-R/FR)的光质条件下生长的拟南芥受热胁迫损伤较轻, 这主要是由于L-R/FR降低了PhyB的活性, 增大了PIFs的丰度(Arico et al., 2019), 表明PhyB介导的光信号影响植物对高温胁迫的适应性.研究发现, PIF4和PIF5可能通过包括激素信号通路在内的多种生物学过程有效促进热胁迫诱导的拟南芥叶片衰老(Li et al., 2021).这种逆境下老叶衰老加速的机制可以清除受损组织, 并促进向幼嫩组织输送营养物质, 提高植物的逆境适应性(Quint et al., 2016). ... The heat shock response: life on the verge of death 1 2010 ... 植物对高温等非生物胁迫响应的分子机制包括感知、信号转导、转录、转录处理、翻译和翻译后蛋白质修饰等多个过程(Zhang et al., 2022).植物感知高温及信号转导对光合作用的稳定至关重要.环境温度升高通过增加质膜的流动性激活Ca2+通道, 促进Ca2+内流至细胞质, 产生胞内信号(Sajid et al., 2018), 并继续通过Ca2+、ROS和不同类型的蛋白激酶等第二信使传递并放大, 引起转录因子的激活和植物激素的级联控制反应(Mittler et al., 2012), 进而激活包括热激蛋白(HSPs)等在内的相对保守的调控网络(Richter et al., 2010), 稳定高温胁迫下叶绿体的发育和PSII的功能(Zhong et al., 2013).而热激因子(heat shock factor, HSF)作为ROS的感受器(Charng et al., 2007), 在调控HSPs表达及参与受损蛋白修复的过程中起关键作用(Qu et al., 2013).位于叶绿体的HSP21被GUN5 (GENOMES UNCOUPLED 5)依赖的逆向信号通路激活, 并通过直接结合D1和D2蛋白等核心亚基稳定高温胁迫下PSII的功能(Chen et al., 2017).此外, 在热适应提高植物高温胁迫耐受性过程中, 植物对热胁迫的记忆依赖于HSPs诱导(Wu et al., 2013)及叶绿体中Ca2+浓度的升高(Pollastri et al., 2021). ... Nonphotochemical chlorophyll fluorescence quenching: mechanism and effectiveness in protecting plants from photodamage 1 2016 ... 研究发现, CEF的重要生理功能是参与非光化学淬灭(NPQ)的诱导、PSI复合体的氧化还原调控以及ATP/NADPH比例的调节(黄伟等, 2012; Suorsa et al., 2016).NPQ是植物进行热耗散的主要机制(Müller et al., 2001; Nishiyama et al., 2001), 依赖于跨膜质子梯度(proton gradient, ΔpH)的高能态荧光淬灭(qE)是NPQ的主要组成部分(Ruban, 2016).而逆境下CEF是叶绿体中ΔpH形成的主要来源(Aihara et al., 2016).高温胁迫下, 植物激活CEF, 通过促进ΔpH使较高的qE热耗散(Sun et al., 2017) (图2).此外, 有证据表明H2O2可能作为环境胁迫和CEF诱导的连接因素, 并在激活CEF过程中起作用(Strand et al., 2015).Ca2+在此过程中也发挥重要作用(Lascano et al., 2003).综合前人的研究结果, 推测高温胁迫主要通过引起过剩光能积累导致ROS产生, ROS进而作为信号分子, 通过Ca2+信号级联反应诱导CEF活性. ... Photosystem II regulation and dynamics of the chloroplast D1 protein in Arabidopsis leaves during photosynthesis and photoinhibition 1 1995 ... 光照不足会导致同化力减弱和光合作用的关键酶未能充分活化进而限制光合作用; 而光强过高会造成植物吸收超过其自身能够利用的光能, 导致过剩激发能的累积, 不可避免地产生ROS, 引发严重的光抑制(Takahashi and Murata, 2008).为了应对时刻变化的光照环境, 植物通过调节光合电子传递产物ATP以及NADPH来适应碳同化的消耗(Foyer et al., 2012).此外, 植物还通过编码2个光系统反应中心亚基的基因快速响应强光(Murchie et al., 2005).高温黑暗对植物PSII活性的伤害已被广泛研究(Li et al., 2009; Xu et al., 2014; Essemine et al., 2016).有趣的是, 研究发现光对高温胁迫下植物光系统具有保护作用(Havaux et al., 1991; Marutani et al., 2012; Buchner et al., 2015).光强直接影响D1蛋白的周转(Tyystj?rvi and Aro, 1996).在一定光强范围内, 随着光强的增加, D1蛋白的周转速度加快, 在中等光强下达到最大值(Russell et al., 1995).Rubisco大、小亚基的含量在黑暗下降低, 而在光下则增高(Demire- vska-Kepova et al., 2005).RCA活性也随着光强的增加而增强(姜振升等, 2010), 从而可能对RCA基因表达进行转录后调控(DeRidder and Salvucci, 2007).研究发现CEF途径受光照影响(Teicher and Scheller, 1998).弱光下CEF的增加对水稻抵御夏季高温逆境具有重要作用(Yamori et al., 2015).这种机制对于探究通过提高植物光合作用抵御高温胁迫具有重要意义. ... SUMOylome profiling reveals a diverse array of nuclear targets modified by the SUMO liga- se SIZ1 during heat stress 1 2018 ... 翻译后修饰是植物快速调节底物蛋白性质和活性的重要机制(Qazi et al., 2019), 也参与逆境胁迫下叶绿体蛋白的调控(Grabsztunowicz et al., 2017).研究发现, SUMO化作为重要的翻译后修饰机制, 能够参与调节植物细胞的热胁迫反应(Rytz et al., 2018).叶绿体是植物细胞中对高温最敏感的细胞器, 为应对高温胁迫, 叶绿体内发生大规模的蛋白质组重塑.叶绿体中的大量蛋白由核基因组编码, 高温导致叶绿体蛋白出现错误定位.SUMO化修饰可以通过蛋白磷酸化正向调节前体蛋白向叶绿体转运的效率, 有助于维持其在叶绿体的定位(Zheng et al., 2022). ... HSP transcript and protein accumulation in brassinosteroid barley mutants acclimated to low and high temperatures 1 2020 ... 油菜素内酯(brassinosteroids, BR)影响植物对各种生物和非生物胁迫的耐受性并调控植物的生长发育.BR在HSP诱导中发挥重要作用(Sadura et al., 2020), 并增强抗氧化酶活性(Cao and Zhao, 2008).油菜素类固醇反应的关键调节因子(BZR1)参与BR介导的植物发育过程, 通过调控活性氧信号代谢通路响应高温胁迫(Yin et al., 2018).研究发现, 高温胁迫下外施BR通过提高植物叶片羧化效率和增强抗氧化系统功能进而缓解光抑制(Ogweno et al., 2008), 并促进气孔导度增大, 维持叶片CO2固定速率(Thus sagunpanit et al., 2015). ... Mechanisms of heat sensing and responses in plants. It is not all about Ca2+ ions 1 2018 ... 植物对高温等非生物胁迫响应的分子机制包括感知、信号转导、转录、转录处理、翻译和翻译后蛋白质修饰等多个过程(Zhang et al., 2022).植物感知高温及信号转导对光合作用的稳定至关重要.环境温度升高通过增加质膜的流动性激活Ca2+通道, 促进Ca2+内流至细胞质, 产生胞内信号(Sajid et al., 2018), 并继续通过Ca2+、ROS和不同类型的蛋白激酶等第二信使传递并放大, 引起转录因子的激活和植物激素的级联控制反应(Mittler et al., 2012), 进而激活包括热激蛋白(HSPs)等在内的相对保守的调控网络(Richter et al., 2010), 稳定高温胁迫下叶绿体的发育和PSII的功能(Zhong et al., 2013).而热激因子(heat shock factor, HSF)作为ROS的感受器(Charng et al., 2007), 在调控HSPs表达及参与受损蛋白修复的过程中起关键作用(Qu et al., 2013).位于叶绿体的HSP21被GUN5 (GENOMES UNCOUPLED 5)依赖的逆向信号通路激活, 并通过直接结合D1和D2蛋白等核心亚基稳定高温胁迫下PSII的功能(Chen et al., 2017).此外, 在热适应提高植物高温胁迫耐受性过程中, 植物对热胁迫的记忆依赖于HSPs诱导(Wu et al., 2013)及叶绿体中Ca2+浓度的升高(Pollastri et al., 2021). ... Beat the heat: plant- and microbe-mediated strategies for crop thermotolerance 1 2022 ... 高温胁迫影响植物各种生理生化过程, 其中植物光合作用的响应取决于胁迫强度和持续时间(Hüve et al., 2011).中度热胁迫(mild heat stress)引起光呼吸增强以及气孔导度降低, 从而使光合作用短暂下降.严重热胁迫(serious heat stress)或热激(heat shock)下, 由于类囊体膜完整性丧失和光系统失活, 引发叶片光合电子传输能力降低(Yamamoto, 2016).如果高温胁迫时间超过植物耐热阈值, 将导致光合机构发生不可逆的损害, 进而对植物光合碳同化过程产生持久影响.因此高温胁迫对植物的伤害及其防御机制研究越来越受到重视(Chen et al., 2022; Shekhawat et al., 2022).在总结前人研究结果的基础上, 笔者对高温胁迫下植物光合作用特性以及提高植物耐高温途径相关研究进行综述, 并展望了高温胁迫下植物光合作用的研究方向. ... Analysis of chlorophyll fluorescence parameters as predictors of biomass accumulation and tolerance to heat and drought stress of wheat (Triticum aestivum) plants 1 2022 ... 在高温胁迫下, 植物采取的光保护机制包括2方面: 避免暴露在强光环境及合理应对过剩光能(Takahashi and Badger, 2011).从整株尺度上, 植物通过树冠内相互遮蔽或调节叶片角度减少受辐射强度, 以及通过蒸腾作用等途径降低叶温(Tserej and Feeley, 2021).由于气孔导度的瞬态增加, 导致蒸腾作用增大, 而蒸腾作用是植物主要降温机制; 此外, 通过调节小气候, 引起同一林分内不同物种甚至同一树冠内不同叶片之间的叶温出现明显差异, 维持大部分叶片的叶温处于光合作用的适宜温度范围内, 避免受极端高温的损害(Fauset et al., 2018).在叶片尺度上避免高温伤害的策略有叶片运动(Kao and Forseth, 1992)和叶绿体运动(Wada et al., 2003), 从而避免叶片和叶绿体受到过强光照, 保护光合机构免受高温伤害; 耐热策略则主要有活性氧清除(Nishiyama et al., 2006)、将捕光复合体吸收的过剩光能以热能的形式耗散(Sherstn- eva et al., 2022)、光合电子传递调节(Jiang et al., 2021)及光呼吸(光下植物吸收O2并释放CO2) (Osei- Bonsu et al., 2021)等. ... Simulated heat waves during maize reproductive stages alter reproductive growth but have no lasting effect when applied during vegetative stages 1 2017 ... 长期在高温环境下生长的植物叶片光合作用的最适温度(Topt)可随环境温度的改变而变化, 表现为高温环境下生长的植物光合作用能适应更高的温度.光合作用能力的可塑性使植物在新的生长温度下更有效地进行光合作用(Yamori et al., 2014).光合作用对高温的可塑性可通过高温驯化获得.在高温条件下, 通过表达热稳定的RCA和/或增加RCA含量来维持Rubisco的高活化状态, 该过程对高温驯化很重要.热激蛋白(HSPs)的诱导是高温驯化的另一个重要过程(Vierling, 1991), 对高温胁迫下蛋白质折叠和组装及蛋白质和膜的稳定性都很重要.植物的高温驯化是长期生长在较高温度下发生的光合适应(Yamori et al., 2014).而短期的高温胁迫响应通常是增加光呼吸和降低净光合速率, 这种影响虽然显著但却是可逆的(Siebers et al., 2017).由于反应较慢或很轻微, 不足以评估植物对高温胁迫的实际适应能力.此外, 最新研究通过遗传工程手段创建了一条全新的蛋白合成途径, 该途径由高温响应启动子驱动细胞核融合基因表达D1蛋白, 建立了植物细胞D1蛋白合成的双途径(天然的叶绿体途径和创建的核途径), 通过增加细胞核源D1合成途径显著增强植物对高温的抗性(Chen et al., 2020). ... Carbon assimilation in crops at high temperatures 1 2019 ... 研究发现, 直接参与光合作用的酶的热不稳定性是温度胁迫下光合速率下降的重要原因(Slattery and Ort, 2019).表1总结了高温胁迫对植物光合作用碳同化过程酶活性的影响.在生化水平上, 植物净光合速率在很大程度上取决于核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)的活化和活性, 以及二磷酸核酮糖(RuBP)的再生(Farquhar et al., 1980).Rubisco是一种双功能酶, 超过35°C的高温胁迫可抑制橡树(Quercus pubescens)叶片Rubisco的初始羧化活力(Haldimann and Feller, 2004).由于Rubisco对CO2和O2的特异性随着温度的升高而降低, 从而降低了体内羧化和氧化反应的比例.因此, 在高温胁迫下Rubisco倾向于氧化反应, 产生更多的2-磷酸乙醇酸, 进入光呼吸途径, 在消耗能量的同时, 也导致光合碳固定的损失(Von Caemmerer, 2020).Rubisco活性受到Rubisco活化酶(RCA)的调节(Bracher et al., 2017).研究表明, RCA具有ATP酶活性, 能够将磷酸糖类抑制物从Rubisco活性位点释放, 从而活化Rubisco (Stotz et al., 2011).RCA活化过程依赖于ATP而受ADP抑制(Kallis et al., 2000).RCA活性易受高温胁迫抑制(Crafts-Brandner and Salvucci, 2000), 其基因表达对短期高温胁迫反应敏感(DeRidder and Salvucci, 2007).因此提高RCA的热稳定性成为缓解高温胁迫对光合能力和作物产量影响的重要手段. ... Photoreceptor phyB involved in Arabidopsis temperature perception and heat-tolerance formation 1 2017 ... 光质主要通过植物感光器对高温胁迫响应起作用.研究表明, 植物体内光、温信号存在分子联系, 其中光敏色素B (PhyB)不仅起光受体的作用, 还具有温度受体的作用.远红光(FR)和适度高温使PhyB失活(Jung et al., 2016; Legris et al., 2017).而拟南芥PhyB突变体的耐热性增强(Song et al., 2017), 表明PhyB在植物适应高温过程中起重要作用.此外, 光敏色素相互作用因子(PIFs)等转录因子在光信号和热形态建成中均发挥作用(Lau et al., 2018).与自然光下生长的拟南芥相比, 在低红光/远红光比率(L-R/FR)的光质条件下生长的拟南芥受热胁迫损伤较轻, 这主要是由于L-R/FR降低了PhyB的活性, 增大了PIFs的丰度(Arico et al., 2019), 表明PhyB介导的光信号影响植物对高温胁迫的适应性.研究发现, PIF4和PIF5可能通过包括激素信号通路在内的多种生物学过程有效促进热胁迫诱导的拟南芥叶片衰老(Li et al., 2021).这种逆境下老叶衰老加速的机制可以清除受损组织, 并促进向幼嫩组织输送营养物质, 提高植物的逆境适应性(Quint et al., 2016). ... Photoinhibition of photosystem I 1 2011 ... 相对PSII而言, PSI对中等程度的高温胁迫不敏感, 特别是在高温强光胁迫下, PSII往往作为胁迫的主要作用位点.研究发现, 高温胁迫提高PSI反应中心P700+的还原速率(Wang et al., 2006), 表明高温会促进PSI的电子传递速率.自然条件下, 植物常处于光强波动环境中, 叶片表面光强的快速增高导致从PSII传递至PSI的电子不能立即被消耗, 造成PSI的过度还原(Yamamoto and Shikanai, 2019), 而此时出现高温胁迫则会引起PSI光抑制(Tan et al., 2020).PSI一旦发生光抑制, 由于恢复时间较长, 往往对光合作用造成更大影响(Sonoike, 2011). ... Structure of green- type Rubisco activase from tobacco 1 2011 ... 研究发现, 直接参与光合作用的酶的热不稳定性是温度胁迫下光合速率下降的重要原因(Slattery and Ort, 2019).表1总结了高温胁迫对植物光合作用碳同化过程酶活性的影响.在生化水平上, 植物净光合速率在很大程度上取决于核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)的活化和活性, 以及二磷酸核酮糖(RuBP)的再生(Farquhar et al., 1980).Rubisco是一种双功能酶, 超过35°C的高温胁迫可抑制橡树(Quercus pubescens)叶片Rubisco的初始羧化活力(Haldimann and Feller, 2004).由于Rubisco对CO2和O2的特异性随着温度的升高而降低, 从而降低了体内羧化和氧化反应的比例.因此, 在高温胁迫下Rubisco倾向于氧化反应, 产生更多的2-磷酸乙醇酸, 进入光呼吸途径, 在消耗能量的同时, 也导致光合碳固定的损失(Von Caemmerer, 2020).Rubisco活性受到Rubisco活化酶(RCA)的调节(Bracher et al., 2017).研究表明, RCA具有ATP酶活性, 能够将磷酸糖类抑制物从Rubisco活性位点释放, 从而活化Rubisco (Stotz et al., 2011).RCA活化过程依赖于ATP而受ADP抑制(Kallis et al., 2000).RCA活性易受高温胁迫抑制(Crafts-Brandner and Salvucci, 2000), 其基因表达对短期高温胁迫反应敏感(DeRidder and Salvucci, 2007).因此提高RCA的热稳定性成为缓解高温胁迫对光合能力和作物产量影响的重要手段. ... Activation of cyclic electron flow by hydrogen peroxide in vivo 1 2015 ... 研究发现, CEF的重要生理功能是参与非光化学淬灭(NPQ)的诱导、PSI复合体的氧化还原调控以及ATP/NADPH比例的调节(黄伟等, 2012; Suorsa et al., 2016).NPQ是植物进行热耗散的主要机制(Müller et al., 2001; Nishiyama et al., 2001), 依赖于跨膜质子梯度(proton gradient, ΔpH)的高能态荧光淬灭(qE)是NPQ的主要组成部分(Ruban, 2016).而逆境下CEF是叶绿体中ΔpH形成的主要来源(Aihara et al., 2016).高温胁迫下, 植物激活CEF, 通过促进ΔpH使较高的qE热耗散(Sun et al., 2017) (图2).此外, 有证据表明H2O2可能作为环境胁迫和CEF诱导的连接因素, 并在激活CEF过程中起作用(Strand et al., 2015).Ca2+在此过程中也发挥重要作用(Lascano et al., 2003).综合前人的研究结果, 推测高温胁迫主要通过引起过剩光能积累导致ROS产生, ROS进而作为信号分子, 通过Ca2+信号级联反应诱导CEF活性. ... Induction of cyclic electron flow around photosystem I du- ring heat stress in grape leaves 1 2017 ... 研究发现, CEF的重要生理功能是参与非光化学淬灭(NPQ)的诱导、PSI复合体的氧化还原调控以及ATP/NADPH比例的调节(黄伟等, 2012; Suorsa et al., 2016).NPQ是植物进行热耗散的主要机制(Müller et al., 2001; Nishiyama et al., 2001), 依赖于跨膜质子梯度(proton gradient, ΔpH)的高能态荧光淬灭(qE)是NPQ的主要组成部分(Ruban, 2016).而逆境下CEF是叶绿体中ΔpH形成的主要来源(Aihara et al., 2016).高温胁迫下, 植物激活CEF, 通过促进ΔpH使较高的qE热耗散(Sun et al., 2017) (图2).此外, 有证据表明H2O2可能作为环境胁迫和CEF诱导的连接因素, 并在激活CEF过程中起作用(Strand et al., 2015).Ca2+在此过程中也发挥重要作用(Lascano et al., 2003).综合前人的研究结果, 推测高温胁迫主要通过引起过剩光能积累导致ROS产生, ROS进而作为信号分子, 通过Ca2+信号级联反应诱导CEF活性. ... PGR5-PGRL1-dependent cyclic electron transport modulates linear electron transport rate in Arabidopsis thaliana 1 2016 ... 研究发现, CEF的重要生理功能是参与非光化学淬灭(NPQ)的诱导、PSI复合体的氧化还原调控以及ATP/NADPH比例的调节(黄伟等, 2012; Suorsa et al., 2016).NPQ是植物进行热耗散的主要机制(Müller et al., 2001; Nishiyama et al., 2001), 依赖于跨膜质子梯度(proton gradient, ΔpH)的高能态荧光淬灭(qE)是NPQ的主要组成部分(Ruban, 2016).而逆境下CEF是叶绿体中ΔpH形成的主要来源(Aihara et al., 2016).高温胁迫下, 植物激活CEF, 通过促进ΔpH使较高的qE热耗散(Sun et al., 2017) (图2).此外, 有证据表明H2O2可能作为环境胁迫和CEF诱导的连接因素, 并在激活CEF过程中起作用(Strand et al., 2015).Ca2+在此过程中也发挥重要作用(Lascano et al., 2003).综合前人的研究结果, 推测高温胁迫主要通过引起过剩光能积累导致ROS产生, ROS进而作为信号分子, 通过Ca2+信号级联反应诱导CEF活性. ... The chilling injury induced by high root temperature in the leaves of rice seedlings 1 2008 ... 除了空气高温对植物光合作用会产生抑制以外, 根区高温对玉米(Zea mays) (Xia et al., 2021)和葡萄(Vitis vinifera) (王浩等, 2022)等叶片的光合特性也具有显著的抑制作用.根区高温一方面通过抑制根系吸收水分和养分影响叶片PSI和PSII的组装(Suzuki et al., 2008); 另一方面通过调节根系呼吸和叶片光合作用之间的平衡改变叶片碳平衡(Atkin et al., 2005).因此, 确定作物特别是设施水培条件下作物生长的最适根区温度, 对于提高生产效率具有重要的指导意义(Yamori et al., 2022). ... Photoprotection in plants: a new light on photosystem II damage 3 2011 ... 植物光系统由光系统I (photosystem I, PSI)和光系统II (PSII)构成.PSII是光合机构中对高温胁迫最敏感的部分之一(Berry and Bjorkman, 1980), 是光抑制的主要发生部位.高温胁迫下, 由于类囊体膜的流动性增强, PSII捕光复合体很容易从类囊体膜上脱落, 从而破坏PSII的完整性, 影响光合电子传递(Mathur et al., 2011).高温胁迫对PSII放氧复合体(oxygen evolving complex, OEC) (Tyystj?rvi, 2012)、反应中心及电子受体侧(Wise et al., 2004)均产生抑制作用, 其抑制程度主要取决于光破坏和光修复的速率.如果PSII的破坏速率超过修复速率, 将导致光能利用效率降低, 造成光抑制(Takahashi and Badger, 2011).高温胁迫下, 当光合机构吸收的过剩激发能不能被及时耗散时, 容易引发活性氧(reactive oxygen species, ROS)累积, 一方面直接破坏PSII反应中心蛋白; 另一方面通过对D1蛋白的从头合成阻碍PSII修复过程, 从而加剧PSII光抑制(Takahashi and Badger, 2011) (图1). ...
本文的其它图/表
-
 图1
高温胁迫诱导植物PSII发生光抑制
图1
高温胁迫诱导植物PSII发生光抑制
高温胁迫导致PSII处的放氧复合体及光合电子传递链失活(1), 或者通过抑制碳固定, 使过剩激发能引发ROS大量积累, ROS一方面直接损伤光合机构组分(2), 另一方面通过抑制D1蛋白的从头合成, 导致D1蛋白的净损失(3), 引起PSII光抑制。D1: D1蛋白; D2: D2蛋白; OEC: 放氧复合体; PSII: 光系统II; ROS: 活性氧
-
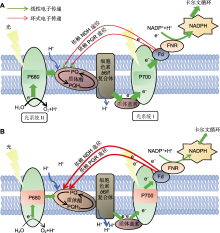 图2
高温胁迫下植物光合电子传递链的变化
图2
高温胁迫下植物光合电子传递链的变化
(A) 光合电子和质子的定向转移依赖于类囊体膜中的关键蛋白复合体; 适宜环境温度下, PSII裂解H2O中的电子主要通过线性电子传递(LEF)至光系统I, 在Fd和FNR的作用下, 将NADP+还原成NADPH, 形成固定CO2所需的还原力; (B) 高温胁迫下, LEF被抑制, PQ库被还原的同时, 依赖NDH及质子梯度调节蛋白/质子梯度调节类似蛋白(PGR5/PGRL1)的环式电子传递(CEF)被激活; LEF的下调可以通过降低传递至光系统I处的电子促进P700的氧化, 而CEF的上调可以通过增加跨膜质子梯度来产生更多的ATP, 也可以通过激发非光化学淬灭来保护光合机构; 因此, 高温胁迫下, LEF和CEF协同作用调节PQ库及P700的氧化还原状态及热耗散, 起到光保护作用。P680和P700分别表示PSII和PSI反应中心叶绿素在可见光谱中的波长吸收峰值。绿色箭头表示LEF转移, 红线表示CEF转移。Fd: 铁氧还蛋白; FNR: 黄素蛋白铁氧还蛋白-NADP还原酶; NADP+: 氧化型烟酰胺腺嘌呤二核苷磷酸; NDH: NA(P)H脱氢酶复合体; PQ: 质体醌; PQH2: 质体醌醇
-
 表1
高温胁迫对植物光合作用碳同化过程酶活性的影响
表1
高温胁迫对植物光合作用碳同化过程酶活性的影响
|

 首页
首页