|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
可变剪切在植物成花转换中的作用
植物学报
2022, 57 (1):
69-79.
DOI: 10.11983/CBB21085
精确调控成花转换, 确保植物在适宜环境下开花, 对于植物的成功繁殖和物种繁衍至关重要。开花由多种分子机制在转录、转录后和蛋白质水平进行调控。可变剪切(AS)是一种普遍的转录后水平调控过程, 可从单个基因产生多个转录本, 从而丰富转录组和蛋白质组的多样性。大量研究表明, 可变剪切在成花转换过程中发挥重要作用。根据发育和环境条件, AS能够影响mRNA的稳定性和/或蛋白亚型的功能, 从而调控开花相关基因的功能转录本和/或功能蛋白水平。揭示成花相关pre-mRNA的AS作用将进一步增进人们对开花相关基因功能以及整个成花转换调控网络的认识。该文归纳了涉及成花转换的AS研究进展, 并针对各个调控途径进行总结, 以期为进一步研究植物AS和成花转换调控机制提供参考。
表1
基因的可变剪切调控成花转换
正文中引用本图/表的段落
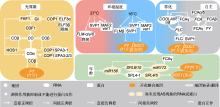
植物进化出复杂的调控网络以感知和整合内源生长信号和外部环境刺激, 确保在合适的条件下开花。在众多环境因素中, 光周期和温度是决定开花时间的主要因素, 涉及光周期途径、春化途径和环境温度途径; 而年龄途径、自主途径和激素途径响应内源生长信号。大量研究表明, AS可通过整合内外源信号而在转录后水平上调控成花转换(表1; 图1)。
在光周期途径中, 拟南芥的CO基因具有核心作用, 其表达在不同水平上受到调控。在转录水平上, CO受CDF1 (CYFLING DOF FACTOR 1)蛋白的抑制, CDF1直接作用于CO启动子以抑制其表达(Seaton et al., 2015)。在光照条件下, FKF1 (FLAVIN-BINDING KELCH REPEAT F-BOX 1)和GI互作形成具有降解CDF1功能的蛋白复合物; 而在黑暗条件下, FKF1与GI则不发生互作, 而且E3泛素连接酶COP1 (CONSTITUTIVE PHOTOMORPHOGENIC 1)与ELF3 (EARLY FLOWERING 3)互作并通过蛋白降解方式影响GI的积累(Jung et al., 2007; Yu et al., 2008; 马朝峰和戴思兰, 2019)。因此, 光照可下调CDF1的蛋白水平, 进而促进CO转录。
在成花转换中, 温度也是关键的调控信号。许多温带植物需要一定时间的冬季低温诱导才能促进春季开花, 这一过程被称为春化(Michaels et al., 2005)。春化作用能够抑制许多开花阻遏基因的表达, 使植物在随后的温暖环境下开花。开花阻遏基因FLC编码MADS-box转录因子, 其是拟南芥和其它相关物种春化途径中的关键调节因子, 通过与SVP蛋白互作而抑制开花整合基因FT和SOC1的表达(Searle et al., 2006)。目前, FLC的AS调控已得到较为系统的研究, 其有义和反义转录本均涉及AS和成花转换。
在调控FT和SOC1表达时, FLC需要与SVP互作形成复合物。SVP同样受到AS调控, 产生常规功能蛋白SVP1和截短蛋白SVP3。其中, 常规功能蛋白能够与FLC及其它相关蛋白互作, 导致其超量表达植株出现晚花和花发育异常的表型; 而截短蛋白缺少互作基序而不能与其它蛋白互作, 因此其超量表达植株不具有相关表型(Severing et al., 2012)。然而, 目前尚未明确SVP的AS是否受温度调控。
植物激素涉及包括成花转换在内的一系列植物生长发育过程。在成花转换调控中, 赤霉素(GAs)起主要作用, 但其它植物激素也均与开花调控相关(Conti, 2017)。大量研究表明, ABA在调节植物开花中发挥多种作用, 并且AS涉及ABA调控的成花转换。ABH1 (ABA hypersensitive 1)参与ABA信号转导, 调节植物对ABA的敏感性; 其编码蛋白是核帽结合蛋白复合物的1个大亚基, 该复合物参与RNA的剪切(Hugouvieux et al., 2001)。而拟南芥abh1突变体对ABA高度敏感并具有早花表型, 而且该突变导致FLC和CO的AS被扰乱(Kuhn et al., 2007)。SKIP (SKI INTERACTING PROTEIN)编码1种含SNW域的剪切因子, 其表达由ABA诱导; SNW域包含1个由5个氨基酸残基组成的SNWKN基序, 为SKIP蛋白与剪接体结合的关键结构域(Lim et al., 2010; Wang et al., 2012)。SKIP调控SEF (SERRATED LEAVES AND EARLY FLOWERING)/SWC6 (SWC6)的AS; 而SEF/SWC6是ATP依赖性染色质重塑复合物的组成部分, 可通过直接结合启动子的近端区域促进FLC表达(Cui et al., 2017; Li et al., 2019)。然而, 目前尚不清楚ABH1和SKIP介导的AS是否响应ABA。
开花整合基因接收各个成花转换途径转导的内外源信号, 以启动下游花分生组织特性基因的表达, 从而精确地控制开花时间。除了各成花转换途径的相关基因, 开花整合基因(如FT和SOC1)也受到AS调控。
本文的其它图/表
|
 首页
首页