
|
|
||
|
濒危植物百山祖冷杉种子发育特征及其胚培养
植物学报
2021, 56 (5):
573-583.
DOI: 10.11983/CBB21096
百山祖冷杉(Abies beshanzuensis)种子萌发率低, 发育特性尚不明确, 极大地限制了种群在原生境的自然更新。该研究以百山祖冷杉不同发育时期的雌球果为材料, 通过研究球果中种子的胚与胚乳数量、重量和显微结构特性, 及测定种子发育关键时期胚乳的初生代谢物, 探究种子发育特征及影响种子发育的主要因素。结果表明, 7月是百山祖冷杉种子发育的关键时期, 此期间胚乳重量快速增加, 胚乳中碳水化合物代谢活动旺盛。解剖学和形态学分析显示, 7月下旬球果中大量种子发育异常。进一步对发育关键时期胚乳中初生代谢物进行分析表明, 海藻糖-6-磷酸含量显著降低。推测在种子发育的关键时期胚乳中海藻糖-6-磷酸降低可能是种子发育异常的重要原因之一。基于种子发育特征, 该研究建立了百山祖冷杉未成熟胚培养技术体系, 并成功获得试管苗。 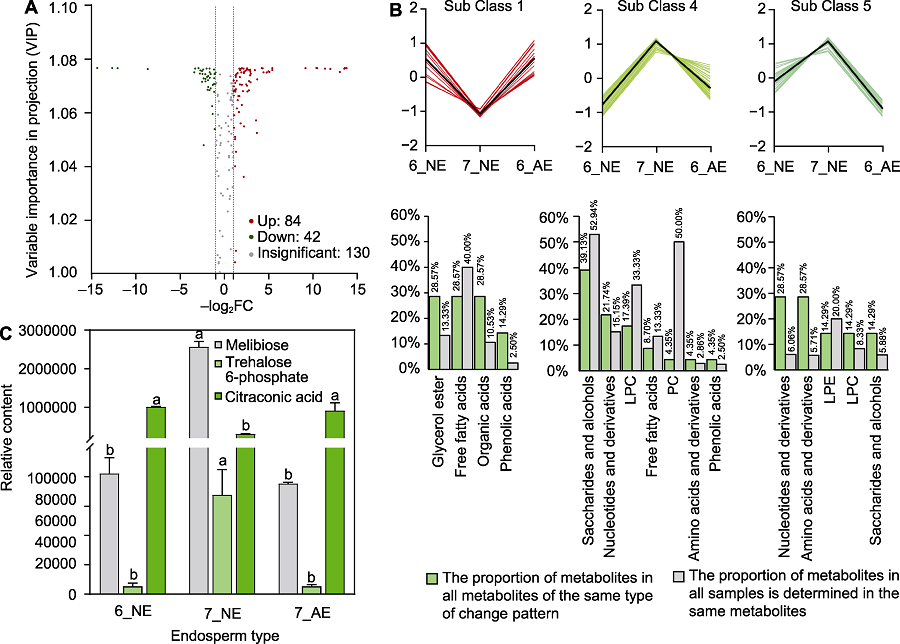 View image in article
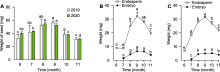
图6
6月和7月百山祖冷杉胚乳的初生代谢物变化
(A) 6月和7月发育正常胚乳的差异代谢物; (B) 第1、4、5类模式中关键差异代谢物数量占每类模式全部代谢物总数的比例与占所有被测样品中同类代谢物总数的比例; (C) 6月和7月百山祖冷杉种子胚乳中的蜜二糖、海藻糖-6-磷酸和甲基马来酸相对含量。6_NE: 6月发育正常的胚乳; 7_NE: 7月发育正常的胚乳; 7_AE: 7月发育异常的胚乳。不同小写字母表示在0.05水平差异显著。FC: 差异倍数值; LPC: 溶血磷脂酰胆碱; LPE: 溶血磷脂酰乙醇胺; PC: 磷脂酰胆碱
正文中引用本图/表的段落
为探究百山祖冷杉种子在胚乳快速生长阶段发育异常种子数迅速增长的原因, 我们利用UPLC-MS/MS检测技术对6月和7月种子中胚乳的初生代谢物类型和含量进行测定。2个时期共检测出256种代谢物, 与6月发育正常的胚乳相比, 7月发育正常的胚乳中有84种代谢物含量显著增加, 42种显著降低(图6A)。
将6月和7月发育正常的胚乳(6_NE和7_NE)及7月发育异常的胚乳(7_AE) 3种类型样本中的代谢物进行K均值(K means)聚类分析, 发现代谢物在不同样本中存在6类不同的变化模式。其中, 第1类代谢物在发育正常的胚乳中的相对含量降低, 而在发育异常的胚乳中其相对含量维持在6月的水平(图6B), 推测此类代谢物含量与正常胚乳在关键时期的发育呈负相关。第4类代谢物在发育正常的胚乳中相对含量升高, 而在7月发育异常的胚乳中相对含量显著低于7月正常的胚乳(图6B)。第5类代谢物在发育正常的胚乳中的变化模式与第4类代谢物相似, 而在7月发育异常的胚乳中的相对含量显著低于6月正常的胚乳(图6B), 推测第4类与第5类代谢物含量与胚乳在关键时期的发育呈正相关。
为明确7月胚乳发育异常的原因, 我们进一步对上述3类模式中的代谢物进行分析, 发现第1类模式中的关键代谢物(7_NE中的含量分别显著低于6_NE和7_AE的代谢物)为甘油酯类、游离脂肪酸类和有机酸类物质, 其中游离脂肪酸类代谢物数量占所有被测样品中同类代谢物总数的40.00% (图6B), 说明7月异常发育胚乳中游离脂肪酸类物质的分解代谢被抑制。第4类模式中的关键代谢物(7_NE中的含量分别显著高于6_NE以及7_AE的代谢物)为糖及醇类、核苷酸及其衍生物类和溶血磷脂酰胆碱(lysophosphati-dylcholine, LPC)类物质, 其中糖及醇类代谢物数量占所有被测样品中同类代谢物总数的52.94% (图6B); 第5类模式中的关键代谢物(7_NE中的含量显著高于6_NE, 且7_AE中的含量显著低于6_NE)为核苷酸及其衍生物、氨基酸及其衍生物和溶血磷脂酰乙醇胺(lysopnosphatidylethanolamine, LPE)类物质(图6B)。上述结果表明, 胚乳在发育关键时期可能需要第4类模式中的糖及醇类物质提供能量, 且第1类模式中游离脂肪酸类物质的积累可能不利于胚乳发育。
利用KEGG数据库对3类模式中的关键代谢物进行注释和代谢途径分析(Kanehisa and Goto, 2000), 发现大量关键差异代谢物参与碳水化合物代谢途径, 包括半乳糖代谢、蔗糖与淀粉代谢及C5-二元酸代谢。对参与碳水化合物代谢途径中关键差异代谢物含量进行分析, 发现7_NE中的蜜二糖和海藻糖-6-磷酸(trehalose 6-phosphate, T6P)含量比6_NE显著增加(图6C), 甲基马来酸含量显著降低(图6C); 与7_NE相比, 7_AE中的海藻糖-6-磷酸含量显著降低(图6C)。已有研究表明, 海藻糖-6-磷酸可通过维持糖代谢平衡进而调控种子发育(Fichtner and Lunn, 2021)。本研究发现, 百山祖冷杉种子发育关键时期, 胚乳中碳水化合物代谢旺盛, 为其快速生长提供了能量来源; 而胚乳中海藻糖-6-磷酸含量降低, 可能会破坏碳水化合物代谢(糖代谢)平衡, 进而导致种子发育异常。
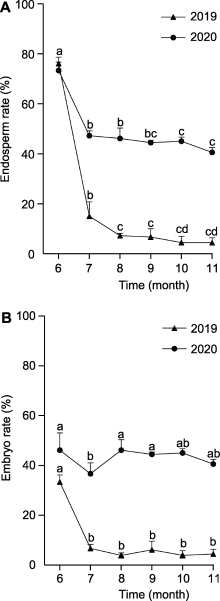
百山祖冷杉种子重量在6-8月迅速增长(图4A), 胚乳中碳水化合物代谢旺盛(图6B)。 然而, 7月开始, 大量种子胚乳中的海藻糖-6-磷酸等物质代谢异常, 发育正常的种子数量骤减(图3)。为解决百山祖冷杉种子自然状态下因代谢异常导致的种子发育问题, 本研究尝试利用胚培养技术对百山祖冷杉的胚进行离体培养(图7A, B)。通过比较百山祖冷杉不同发育时期种子中胚培养的萌发率, 发现胚的萌发率从6-11月呈先上升后下降的趋势。其中, 8月和9月的萌发率最高, 分别为86.67%和93.65% (图7C), 高于7月胚的萌发率(54.17%)。6月的胚处于发育早期, 尚未形成完整植株。此外, 10月开始, 胚的萌发率逐渐降低, 根据对种子的显微观察, 此时胚已分化形成子叶, 接近成熟, 胚萌发率低的原因可能是种子将进入休眠状态, 大量抑制胚萌发物质的逐渐积累。
本文的其它图/表
|
 首页
首页

{kind=link}