|
|
||||||||||||||||||||||||||||||||||||
|
植物微管骨架参与下胚轴伸长调节机制研究进展
植物学报
2021, 56 (3):
363-371.
DOI: 10.11983/CBB20170
微管作为细胞骨架的重要成员, 在植物生长发育过程中起重要作用。下胚轴作为研究细胞伸长的模式系统之一, 其伸长受到多种信号的调节。该文综述了微管骨架在响应环境和生长发育信号调节下胚轴伸长过程中的作用及机制, 旨在帮助读者深入理解微管骨架响应上游信号在植物下胚轴伸长中的作用机理。
表1
参与调节下胚轴伸长的微管相关蛋白
正文中引用本图/表的段落
光是下胚轴伸长的主要调控因子之一。黑暗条件下, 植株下胚轴表现为黄化生长且快速伸长, 见光后下胚轴的伸长速率下降, 说明光对下胚轴伸长有强烈的抑制作用。Le等(2015)研究表明, 在快速伸长的下胚轴细胞内, 周质微管垂直于伸长轴, 表现为横向排列方式, 而在伸长缓慢或停止伸长的下胚轴细胞内, 周质微管表现为斜向或纵向排列; 快速伸长生长的黄化下胚轴见光后, 周质微管由横向平行排列变为斜向或纵向排列, 进而有利于抑制下胚轴的伸长。对光信号调节微管重排的研究发现, 光信号通过改变下胚轴细胞中周质微管的动态及转换能力调控其排列方式, 进而调节下胚轴的伸长。下胚轴细胞在快速伸长之前, 细胞内的微管会形成一种具有双极性的纵向列阵, 并转换为放射状的星状微管列阵, 之后开始进入快速伸长生长。在快速伸长的植株细胞内, 微管的聚合和重排速率都更快(Sambade et al., 2012)。此外, 蓝光可在15分钟内诱导拟南芥下胚轴细胞中横向排列的周质微管完成90度重排。具体机理是: 蓝光照射后微管切割蛋白katanin切割交叉部位的微管产生新末端, 微管正端结合蛋白CLASP (cytoplasmic linker protein-associated protein)稳定新形成的微管末端, 使新末端在伸长过程中改变微管的排列方式为纵向排列(Lindeboom et al., 2013; Wang et al., 2017; Lindeboom et al., 2019) (表1)。该研究结果为揭示光诱导微管重排的分子机理提供了重要线索。
此外, 还发现一些参与光信号调节微管重排的微管相关蛋白在下胚轴伸长中起重要作用, 但目前仍不清楚光信号如何调控这些微管相关蛋白。微管去稳定蛋白25 (microtubule destabilizing protein25, MDP 25)在光抑制下胚轴伸长中起重要作用。光信号引起下胚轴细胞中钙离子浓度增加, 使MDP25蛋白从质膜上脱离并进入细胞质, 通过调节周质微管从横向变为斜向或纵向排列, 在下胚轴由黑暗转移至光下后抑制光下下胚轴伸长过程中起重要作用(Li et al., 2011)。Liu等(2013)发现了1个新的在光抑制下胚轴伸长过程中起正调控作用的微管相关蛋白WDL3 (wave-dampened2-like3)。黑暗条件下, WDL3被26S蛋白酶体降解, 无法抑制下胚轴伸长, 而当黄化苗被转移至光下后, WDL3蛋白能稳定表达, 并通过调节下胚轴细胞内周质微管的组织动态抑制下胚轴伸长。Lian等(2017)研究发现, 黑暗条件下, 植物光形态建成的核心调控因子E3泛素连接酶COP1 (constitutive photomorphogenic1)可直接与WDL3结合并使其降解, 该研究揭示了光信号通过直接调控微管相关蛋白水平来调控周质微管重排的新机制。此外, 还有一些微管相关蛋白只特异性地在光下或黑暗下调节下胚轴的伸长。例如, 微管去稳定蛋白MDP60 (microtu bule destabilizing protein60)和微管稳定蛋白SPR1 (SPIRAL1)。MDP60主要在下胚轴表达, 通过改变微管组织和动态来调节下胚轴伸长, 光通过其信号通路的核心转录因子PIF3 (phytochrome-interacting fac-tor 3)负调控MDP60的表达。PIF3作为乙烯促进光下下胚轴伸长的下游关键调节因子, 其蛋白水平受26S蛋白酶体对光信号响应的调节。在光介导的下胚轴伸长过程中, PIF3与MDP60启动子特异性结合, 从而提高MDP60的表达水平, MDP60通过改变微管排列促进下胚轴伸长(Ma et al., 2018)。微管正端结合蛋白SPR1促进黑暗下生长的下胚轴细胞伸长, 在黑暗下生长的下胚轴中可检测到SPR1的表达, 而在光下生长的下胚轴中未检测到SPR1的表达, 表明光影响SPR1表达(Nakajima et al., 2004, 2006; Wang and Mao, 2019)。但尚不清楚响应光调节SPR1表达的上游转录因子, 具体机理有待后续深入研究。
生长素促进下胚轴伸长可通过调节周质微管为横向排列实现。植株体内缺少生长素会抑制其生长, 细胞内的微管以纵向排列为主, 纵向排列的微管可通过外源添加生长素改变为横向排列, 且这种转变具有剂量依赖效应(Fischer and Schopfer, 1997)。关于生长素信号调控微管骨架排列方式分子机制的研究, 从发现生长素能够调节下胚轴细胞内微管的重新排列就已开始。Chen等(2014)研究表明, 生长素通过生长素结合蛋白1 (auxin binding protein 1, ABP1)调节ROP (Rho of plants) GTPase、ROP的互作蛋白RIC1 (ROP-interactive CRIB motif-containing protein 1)和微管切割蛋白katanin来调控下胚轴细胞内的微管骨架重排。True和Shaw (2020)发现, 外源生长素诱导下胚轴细胞周质微管重排需要转运抑制因子/生长素F-box (transport inhibitor 1/auxin F-box, TIR1/AFB)转录途径。但是关于生长素促进下胚轴伸长与调控微管重排之间的关系尚存在不同观点, 有研究者认为, 下胚轴细胞内周质微管重排由生长本身引起, 而不依赖于生长素(Adamowski et al., 2019)。目前, 在生长素调控下胚轴伸长过程中, 并未发现受生长素信号直接调控的微管蛋白或微管相关蛋白, 对于微管骨架响应生长素信号调节下胚轴伸长的机理也需深入研究。
乙烯是一种气体激素, 在调控植物下胚轴伸长中起重要作用(Smalle et al., 1997; Zhong et al., 2012)。乙烯可根据光照条件促进或抑制拟南芥下胚轴伸长(Ecker et al.,1995; Smalle et al., 1997)。光照条件下, 乙烯或者其前体ACC (1-aminocyclopropane-1-car-boxylic acid)处理可促进下胚轴伸长; 黑暗条件下, 乙烯则抑制下胚轴伸长(Zhong et al., 2012; Yu et al., 2013)。乙烯通过转录因子EIN3激活PIF3依赖的生长促进途径及乙烯响应因子1 (ethylene response fac-tor 1, ERF1)介导的生长抑制途径来调控下胚轴生长。在光下, 乙烯通过其信号途径关键转录因子EIN3直接结合PIF3的启动子区激活其表达, 从而促进光下下胚轴伸长; 黑暗条件下, 乙烯可诱导抑制伸长的ERF1蛋白积累, 进而抑制黄化下胚轴伸长(Zhong et al., 2012)。此外, 乙烯还可通过COP1介导的HY5 (hypocotyl 5)蛋白降解来促进光下下胚轴的伸长(Yu et al., 2013)。
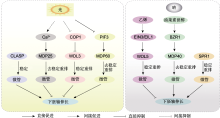
COP1: 持续光形态建成1; EIN3/EIL1: 乙烯不敏感3/乙烯不敏感3类似1. CLASP、MDP25、WDL3、MDP60、SPR1、WDL5、MDP40、PIF3、EIN3和BZR1同
油菜素甾醇(brassinosteroids, BRs)是一种重要的调节植物生长发育的植物激素(Clouse, 2011; Ye et al., 2011)。BRs通过受体激酶BRI1 (brinsensitive1)以及特定的信号转导途径激活2个关键转录因子BZR1 (brassinazole-resistant 1)和BES1/BZR2 (brinsensi-tive1-EMS-suppressor1/brinsensitive2)来起作用(Li et al., 2010; Kim and Wang, 2010; Clouse, 2011; Gudesblat and Russinova, 2011)。许多油菜素甾醇缺陷及不敏感突变体的黄化下胚轴变短且光下生长的植株矮小。例如, 油菜素甾醇合成缺失突变体det2 (de-etiolated-2)在黑暗下生长时子叶张开且下胚轴伸长受到抑制(Chory et al., 1991; Wang et al., 2001); 油菜素甾醇受体BRI1的无效突变体bri1-116植株也表现出黄化下胚轴短的表型; 油菜素甾醇信号途径转录因子BZR1的激活标签突变体bzr1-1D则表现出黄化下胚轴长且弯曲的表型; 油菜素甾醇信号途径的负调控因子BIN2的显性突变体bin2-1植株黄化下胚轴伸长也受到抑制; 油菜素甾醇信号通路中的上游组分BSKs和BSU1通过改变转录因子BZR1的磷酸化状态调控下胚轴的伸长(Tang et al., 2008; Kim et al., 2010; Gudesblat and Russinova, 2011)。上述结果表明, BRs在调控下胚轴伸长方面起重要作用。
微管骨架是细胞骨架的重要成员之一, 参与多种细胞学过程。下胚轴作为研究植物细胞伸长的模式系统, 微管的组织动态变化会影响下胚轴的伸长。同时, 下胚轴伸长受到多种内部和外部信号调控, 这些信号在调控下胚轴伸长过程中均伴随着周质微管组织排列方式的变化, 但微管骨架响应各种内部及外部信号参与调节下胚轴伸长的机理, 以及微管在信号间互相促进或拮抗调控下胚轴伸长中的作用等问题目前仅进行了初步研究, 还有很多科学问题尚待进一步探索。我们对参与下胚轴伸长调节的微管相关蛋白进行了总结(表1), 并对这些微管蛋白如何响应上游信号参与调节下胚轴伸长进行了归纳(图1)。农业生产上, 种子在土壤中萌发后, 胚轴快速伸长以破土而出, 使子叶见光, 进行光合作用。因此, 阐明微管响应各种内部及外部信号转导途径, 调节下胚轴伸长的机理对于解析下胚轴伸长调节的分子机制和农业生产上提高种子萌发率均具有重要意义。
CLASP: Cytoplasmic linker protein-associated protein; MDP25: Microtubule destabilizing protein 25; WDL3: Wave-dampened 2 like 3; MDP60: Microtubule destabilizing protein 60; SPR1: Spiral 1; WDL5: Wave-dampened 2 like 5; MDP40: Microtubule destabilizing protein 40; PIF3: Phytochrome-interacting factor 3; EIN3: Ethylene-insensitive 3; BZR1: Brassinazole-resistant 1 ... Disorganization of cortical microtubules stimulates tangential expansion and reduces the uniformity of cellulose microfibril alignment among cells in the root of Ara- bidopsis 1 2004 ... 微管在植物多种生理活动中起重要作用, 如维持细胞形态、控制细胞极性生长、调控细胞有丝分裂与细胞分化以及参与囊泡运输和信号转导( Insensitivity to ethylene conferred by a dominant mutation in Arabidopsis thaliana 1988 Inhibition of cell expansion by rapid ABP1-mediated auxin effect on microtubules 1 2014 ... 生长素促进下胚轴伸长可通过调节周质微管为横向排列实现.植株体内缺少生长素会抑制其生长, 细胞内的微管以纵向排列为主, 纵向排列的微管可通过外源添加生长素改变为横向排列, 且这种转变具有剂量依赖效应( Phenotypic and genetic analysis of det2, a new mutant that affects light- regulated seedling development in Arabidopsis 1 1991 ... 油菜素甾醇(brassinosteroids, BRs)是一种重要的调节植物生长发育的植物激素( Brassinosteroid signal transduction: from receptor kinase activation to transcriptional networks regulating plant development 2 2011 ... 油菜素甾醇(brassinosteroids, BRs)是一种重要的调节植物生长发育的植物激素(
CLASP: Cytoplasmic linker protein-associated protein; MDP25: Microtubule destabilizing protein 25; WDL3: Wave-dampened 2 like 3; MDP60: Microtubule destabilizing protein 60; SPR1: Spiral 1; WDL5: Wave-dampened 2 like 5; MDP40: Microtubule destabilizing protein 40; PIF3: Phytochrome-interacting factor 3; EIN3: Ethylene-insensitive 3; BZR1: Brassinazole-resistant 1 ... Microtubules and the shape of plants to come 1 2004 ... 此外, 还发现一些参与光信号调节微管重排的微管相关蛋白在下胚轴伸长中起重要作用, 但目前仍不清楚光信号如何调控这些微管相关蛋白.微管去稳定蛋白25 (microtubule destabilizing protein25, MDP 25)在光抑制下胚轴伸长中起重要作用.光信号引起下胚轴细胞中钙离子浓度增加, 使MDP25蛋白从质膜上脱离并进入细胞质, 通过调节周质微管从横向变为斜向或纵向排列, 在下胚轴由黑暗转移至光下后抑制光下下胚轴伸长过程中起重要作用( Coordinated regulation of hypocotyl cell elongation by light and ethy- lene through a microtubule destabilizing protein 2 2018 ... 微管的基本组成单位是由α-微管蛋白和β-微管蛋白构成的微管蛋白异二聚体, 微管蛋白异二聚体首尾相连并线性排列为1根原纤丝, 13根原纤丝平行排列构成中空的管状结构即为微管.植物细胞中的微管始终处于高度动态状态, 描述微管动态特性的模型有2个: (1) 动态不稳定模型.该模型认为单根微管始终处于动态不稳定状态, 单根微管的两端以一定的“收缩速率”解聚或者以一定的“生长速率”聚合, 整体的微管解聚量和聚合量相对平衡, 因此群体微管处于稳定状态(
本文的其它图/表
|
 首页
首页