

 首页
首页
1 材料与方法
1.1 植物材料
1.2 实验方法
1.2.1 牡丹TCP家族成员鉴定、理化性质分析及系统进化树构建
1.2.2 牡丹TCP蛋白二级结构及保守域三级结构预测
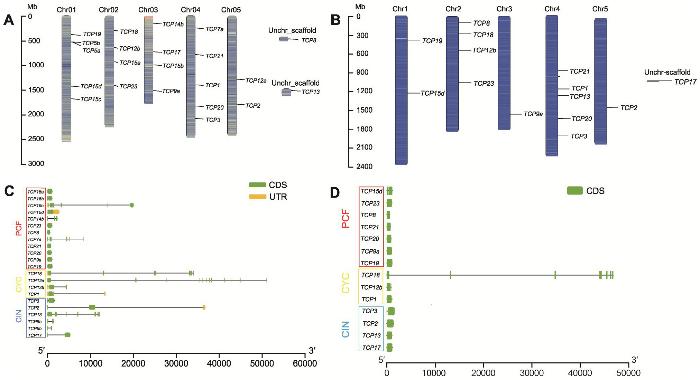
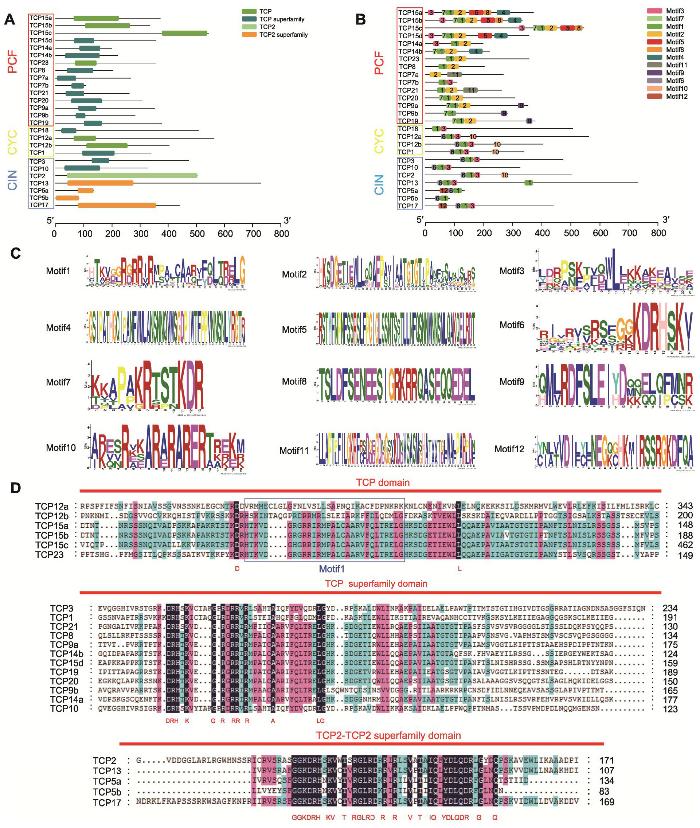
1.2.3 牡丹TCP基因结构、染色体定位及保守基序分析
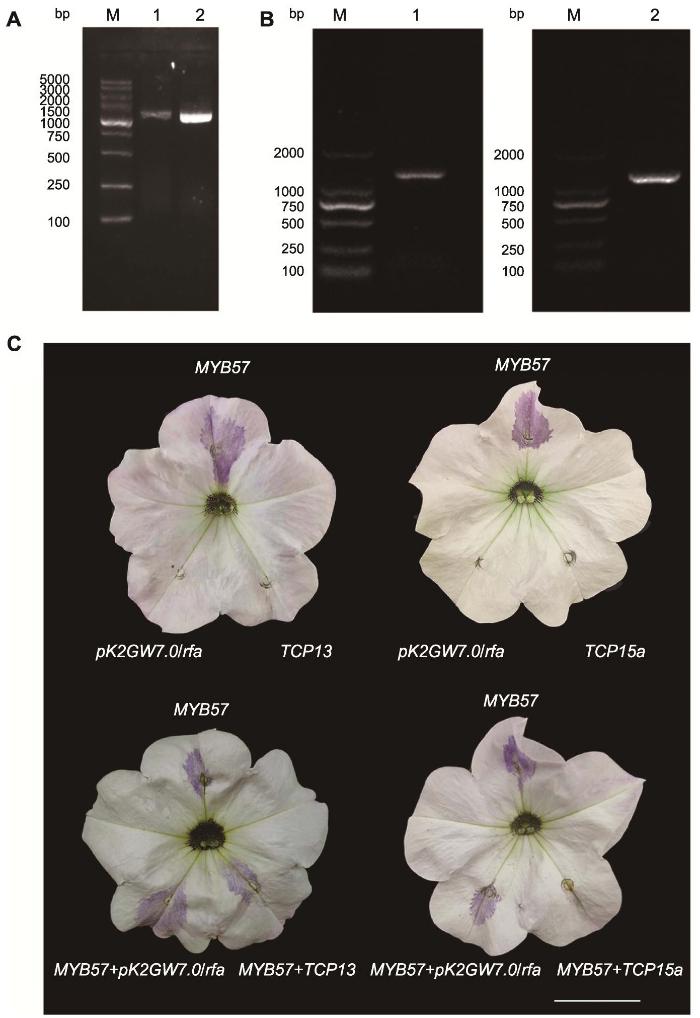
1.2.4 牡丹TCP13和TCP15a基因克隆及载体构建
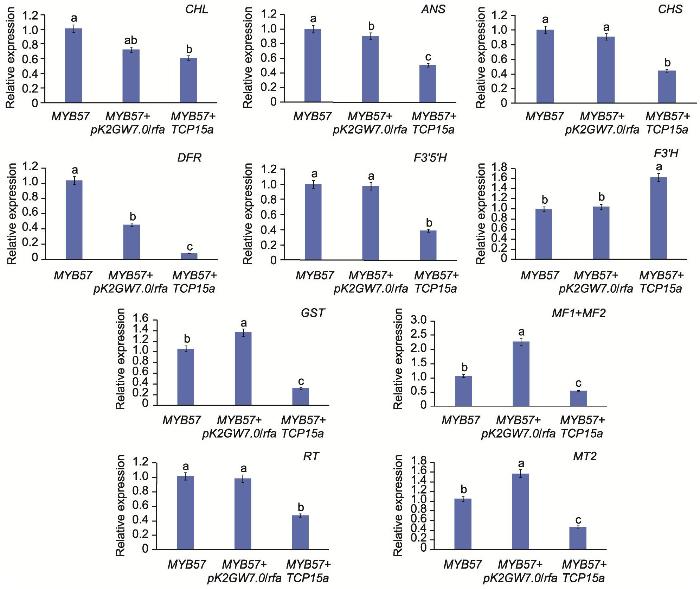
1.2.5 牡丹TCP13和TCP15a瞬时表达及荧光定量分析
表1 qRT-PCR所用引物Table. 1 Primers for qRT-PCR testing |
| Gene | Forward primer sequence(5′–3′) | Reverse primer sequence(5′–3′) |
|---|---|---|
| CHSa | CAACTAGTGGTGTGGACATGCC | CACCAGCAAAGCAACCTTGTT |
| CHI | TCTCCTCCAGTGTCCGTTACTAAA | ACTTCCCTTCTATCTCCAGACCTC |
| F3′H | AGCTGGACGTAGGATTTGTG | ATGGATCAGCCCGTTGTAAG |
| F3′5′H | ATAGGCGTTTACTCGAATCCG | TGGTGTAGAAGGGTGTTTTCG |
| DFR | CCCCTAGTTTAATCACTGCCC | GACCATCTTAGCCACATCGTAG |
| ANS | TCTTCCATTGTGCTTTCCCTG | GTTGCTGGAGTGTAGTCAGTAG |
| RT | CAGGGCTTCCTTTCTTTCTTGTCT | TCCCCTTGTCTTTCACTCTTTCC |
| MT2 | GCAGCGAGCTTTATGGGTCTT | TGAGCTTTAGGAACATCGATAGAAAC |
| MF1+MF2 | CTTTTGGCTACTGCTCTTGCATT | CTCTGTCCGGATCAATCGCTAT |
| GST | GGTGACCAAGAGAAAGTGTTTGC | GCTGGGATAGACACTGCTTCA |
| actin | AGCCAACAGAGAGAAGATGACCCA | ACACCATCACCAGAGTCCAACACA |
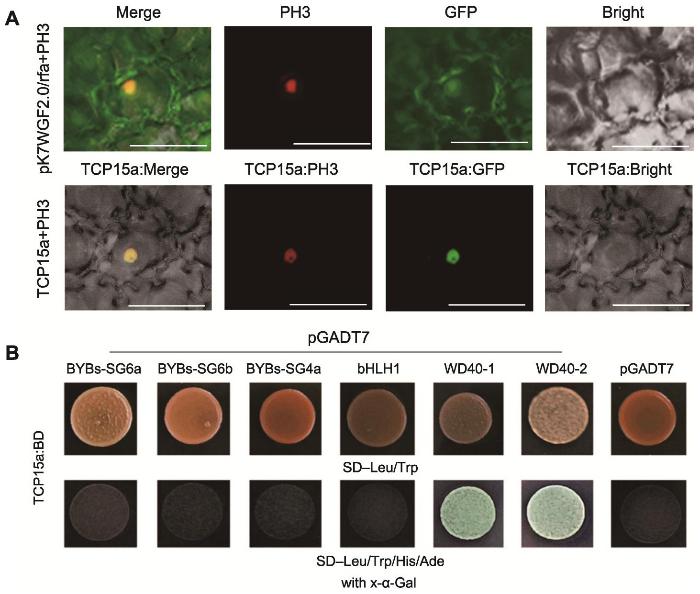
1.2.6 牡丹TCP15a亚细胞定位
1.2.7 酵母双杂交
2 结果与分析
2.1 牡丹TCP基因家族成员系统进化分析、分类及鉴定
表2 牡丹TCP家族成员的理化性质分析Table 2 Analysis of physical and chemical properties of TCP family members in tree peony |
| Gene name | Gene ID | Gene ID | Gene ID | Protein length(aa) | pI | Molecular weight(kDa) | Conserved domains |
|---|---|---|---|---|---|---|---|
| TCP1 | Pos.gene11379 | PL-2G122130.1 | Psu.T.00000087.1 | 339,339,307 | 9.02,8.92,9.59 | 37.75,37.89,33.98 | TCP superfamily |
| TCP2 | Pos.gene53232 | PL-4G132790.1 | Psu.T.00000478.1 | 503,465,464 | 9.02,7.16,7.14 | 54.83,50.50,50.20 | TCP2 |
| TCP3 | Pos.gene72619 | PL-2G273790.1 | Psu.T.00034421.1 | 473,473,530 | 5.83,5.78,7.2 | 51.69,51.73,58.39 | TCP superfamily |
| TCP5a | Pos.gene3141 | / | / | 134,/,/ | 9.51,/,/ | 15.49,/,/ | TCP2 superfamily |
| TCP5b | Pos.gene3142 | / | / | 83,/,/ | 10.06,/,/ | 9.79,/,/ | TCP2 superfamily |
| TCP7a | Pos.gene49570 | / | / | 267,/,/ | 9.64,/,/ | 29.89,/,/ | TCP superfamily |
| TCP7b | / | / | Psu.T.00012308.1 | /,/,107 | /,/,9.88 | /,/,12.16 | TCP superfamily |
| TCP8 | Pos.gene73497 | PL-5G111400.1 | / | 203,217,/ | 8.33,7.66,/ | 21.94,23.23,/ | TCP superfamily |
| TCP9a | Pos.gene42145 | PL-3G121530.1 | / | 352,346,/ | 7.84,8.55,/ | 37.60,37.16,/ | TCP superfamily |
| TCP9b | / | / | Psu.T.00008119.1 | /,/,282 | /,/,9.37 | /,/,29.99 | TCP superfamily |
| TCP10 | / | / | Psu.T.00004485.1 | /,/,325 | /,/,6.89 | /,/,36.38 | TCP superfamily |
| TCP12a | Pos.gene45291 | / | / | 562,/,/ | 9.38,/,/ | 65.01,/,/ | TCP |
| TCP12b | Pos.gene7348 | PL-5G147810.1 | / | 403,270,/ | 8.08,9.35,/ | 45.22,30.17,/ | TCP |
| TCP13 | Pos.gene72744 | PL-2G116620.1 | / | 729,357,/ | 6.25,6.9,/ | 80.39,39.63,/ | TCP2 superfamily |
| TCP14a | / | / | Psu.T.00014829.1 | /,/,198 | /,/,10.12 | /,/,22.30 | TCP superfamily |
| TCP14b | Pos.gene82182 | / | / | 220,/,/ | 9.78,/,/ | 24.15,/,/ | TCP superfamily |
| TCP15a | Pos.gene36892 | / | Psu.T.00029341.1 | 371,/,371 | 6.67,/,6.67 | 39.83,/,39.83 | TCP |
| TCP15b | Pos.gene43777 | / | / | 333,/,/ | 9.62,/,/ | 37.23,/,/ | TCP |
| TCP15c | Pos.gene61444 | / | Psu.T.00005566.1 | 543,/,128 | 5.88,/,11.2 | 60.73,/,14.38 | TCP |
| TCP15d | Pos.gene27661 | PL-1G208530.1 | / | 356,354,/ | 7.82,7.82,/ | 38.31,38.00,/ | TCP superfamily |
| TCP17 | Pos.gene78687 | PL-UG256570.1 | / | 440,378,/ | 9.12,9.36,/ | 49.28,42.08,/ | TCP2 superfamily |
| TCP18 | Pos.gene72830 | PL-5G126440.1 | Psu.T.00026341.1 | 507,701,200 | 6.29,7.55,9.6 | 55.75,78.12,21.96 | TCP superfamily |
| TCP19 | Pos.gene75864 | PL-1G133380.1 | / | 377,377,/ | 5.26,5.26,/ | 39.82,39.82,/ | TCP superfamily |
| TCP20 | Pos.gene22067 | PL-2G301650.1 | Psu.T.00007142.1 | 307,306,309 | 8.68,8.68,8.68 | 32.98,32.88,33.24 | TCP superfamily |
| TCP21 | pos.gene79130 | PL-2G150160.1 | Psu.T.00026535.1 | 261,261,261 | 9.45,9.45,9.45 | 27.42,27.36,27.42 | TCP superfamily |
| TCP23 | Pos.gene34937 | PL-5G204500.1 | / | 356,356,/ | 7.91,7.91,/ | 37.92,37.91,/ | TCP |
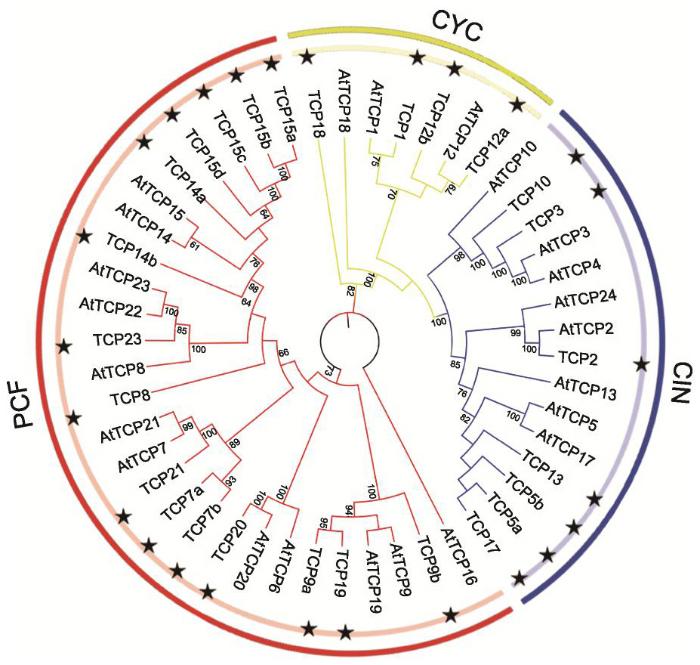
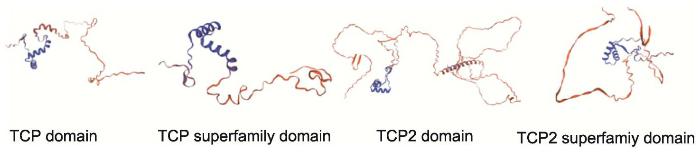
2.2 TCP蛋白二级结构分析及保守域三级结构预测
表3 牡丹TCP基因家族的二级结构分析Table 3 Second structure analysis of the TCP gene family in tree peony |
| Gene name | Alpha helix(proportion) | Extended strand(proportion) | Beta turn(proportion) | Random coil(proportion) | Subgroup |
|---|---|---|---|---|---|
| TCP15a | 33(8.89%) | 20(5.39%) | 5(1.35%) | 313(84.37%) | PCF |
| TCP15b | 26(7.81%) | 17(5.11%) | 3(0.90%) | 287(86.19%) | |
| TCP15c | 200(36.83%) | 94(17.31%) | 25(4.60%) | 224(41.25%) | |
| TCP15d | 41(11.52%) | 12(3.37%) | 5(1.40%) | 298(83.71%) | |
| TCP14a | 26(13.13%) | 9(4.55%) | 2(1.01%) | 161(81.31%) | |
| TCP14b | 26(11.82%) | 12(5.45%) | 4(1.82%) | 178(80.91%) | |
| TCP23 | 37(10.39%) | 15(4.21%) | 8(2.25%) | 296(83.15%) | |
| TCP8 | 53(26.11%) | 7(3.45%) | 3(1.48%) | 140(68.97%) | |
| TCP7a | 45(16.85%) | 29(10.86%) | 10(3.75%) | 183(68.54%) | |
| TCP7b | 25(23.36%) | 8(7.48%) | 3(2.80%) | 71(66.36%) | |
| TCP21 | 38(14.56%) | 11(4.21%) | 2(0.77%) | 210(80.46%) | |
| TCP20 | 38(12.38%) | 8(2.61%) | 5(1.63%) | 256(83.39%) | |
| TCP9a | 33(9.38%) | 24(6.82%) | 6(1.70%) | 289(82.10%) | |
| TCP9b | 19(6.74%) | 19(6.74%) | 6(2.13%) | 238(84.40%) | |
| TCP19 | 44(11.67%) | 19(5.04%) | 7(1.86%) | 307(81.43%) | |
| TCP18 | 143(28.21%) | 73(14.40%) | 20(3.94%) | 271(53.45%) | CYC |
| TCP12a | 193(34.34%) | 58(10.32%) | 29(5.16%) | 282(50.18%) | |
| TCP12b | 92(22.83%) | 21(5.21%) | 4(0.99%) | 286(70.97%) | |
| TCP1 | 85(25.07%) | 9(2.65%) | 1(0.29%) | 244(71.98%) | |
| TCP3 | 43(9.09%) | 16(3.38%) | 2(0.42%) | 412(87.10%) | CIN |
| TCP10 | 42(12.92%) | 24(7.38%) | 2(0.62%) | 257(79.08%) | |
| TCP2 | 77(15.31%) | 29(5.77%) | 5(0.99%) | 392(77.93%) | |
| TCP13 | 151(20.71%) | 99(13.58%) | 27(3.70%) | 452(62.00%) | |
| TCP5a | 17(12.69%) | 8(5.97%) | 1(0.75%) | 108(80.60%) | |
| TCP5b | 19(22.89%) | 8(9.64%) | 2(2.41%) | 54(65.06%) | |
| TCP17 | 57(12.95%) | 30(6.82%) | 2(0.45%) | 351(79.77%) |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}