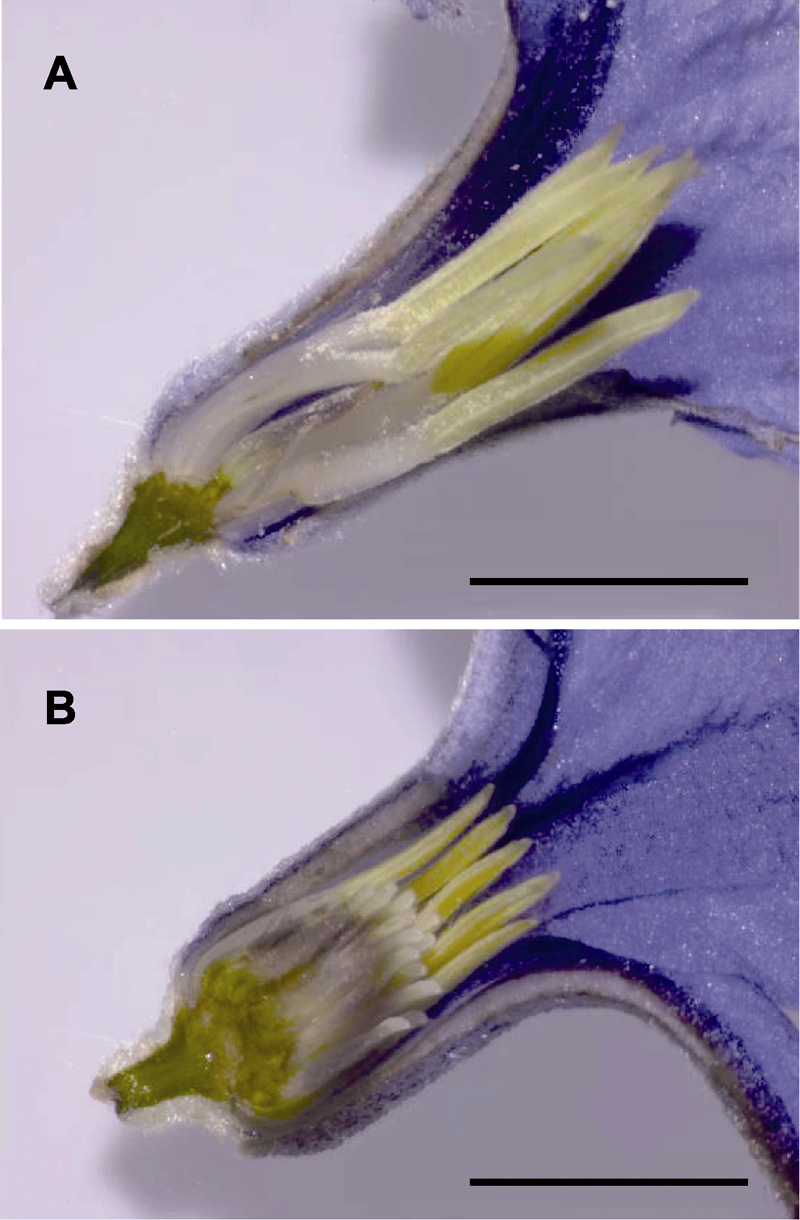
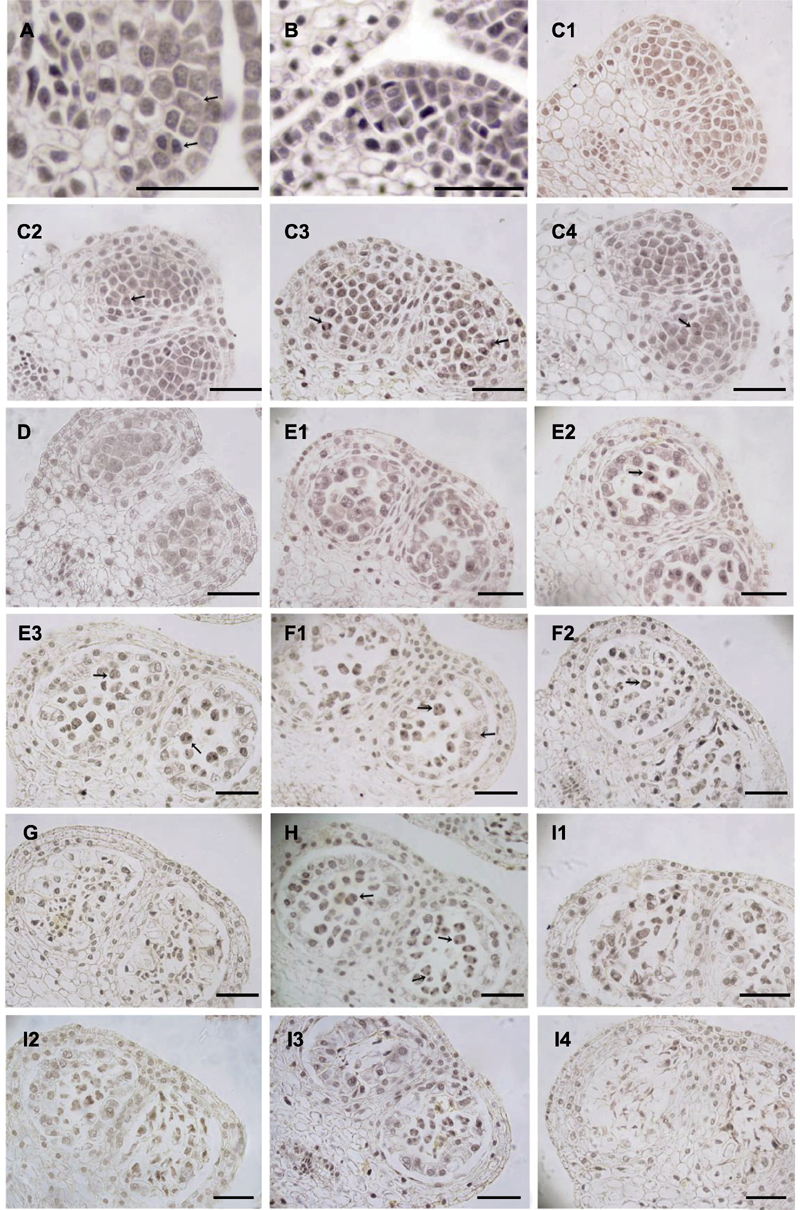
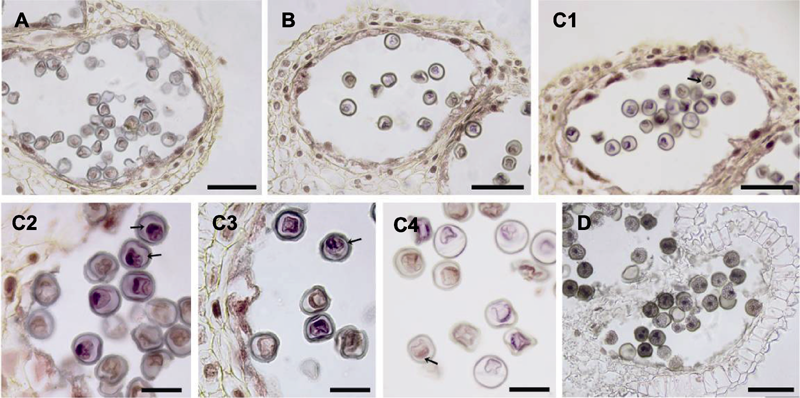
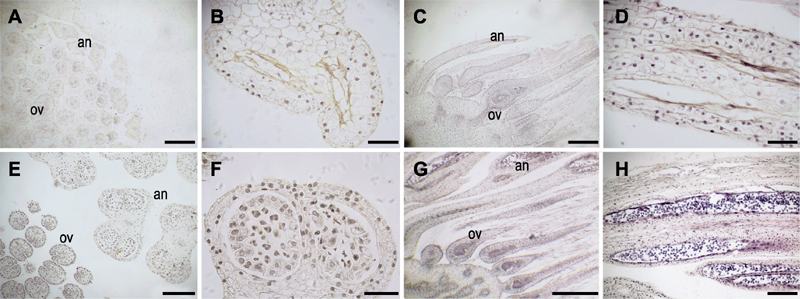
| 1 |
管开云, 李志坚, 李景秀, 匡建 (2002). 铁线莲属植物的引种栽培研究初报. 植物分类与资源学报 24, 392-396.
|
| 2 |
李杨, 张伟, 赫雪锋, 汤海峰 (2018). 铁线莲属植物的化学成分及药理作用研究进展. 中南药学 16, 355-362.
|
| 3 |
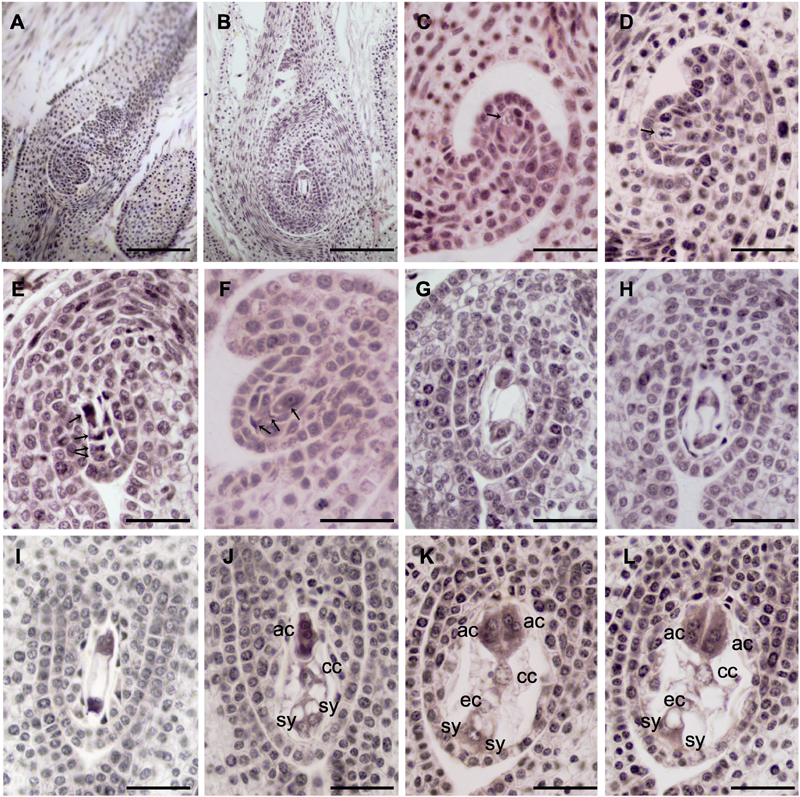
刘萍 (2006). 铁筷子(毛茛科) 的形态、结构、发育及系统位置的研究. 硕士论文. 西安: 陕西师范大学. pp. 15-35.
|
| 4 |
刘庆超, 王奎玲, 卢婉佩, 刘庆华 (2014). 我国铁线莲属植物资源研究进展. 植物遗传资源学报 15, 483-490, 497.
|
| 5 |
全雪丽, 刘继生, 吴松权 (2012). 棉团铁线莲小孢子发生和雄配子体的形成. 辽宁林业科技(1), 9-10, 32.
|
| 6 |
宋春慧 (2006). 黄三七属(毛茛科) 植物的形态、结构、发育及系统位置研究. 硕士论文. 西安: 陕西师范大学. pp. 25.
|
| 7 |
宋萍 (2007). 金莲花属和驴蹄草属(毛茛科)的比较形态学研究——基于形态、结构、花发育及胚胎学的证据. 硕士论文. 西安: 陕西师范大学. pp. 11-18.
|
| 8 |
孙诚, 谢磊, 李良千 (2007). 铁线莲属尾叶铁线莲组(毛茛科)基于形态学证据的分支系统学. 植物学通报 24, 87-98.
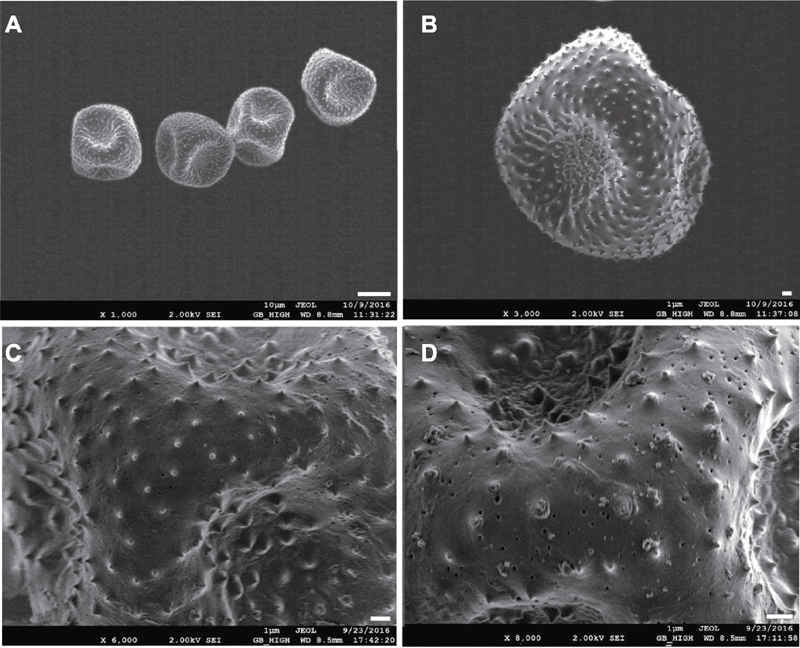
|
| 9 |
王玖瑞, 刘玲, 刘孟军, 周俊义 (2006). 枣树雄性不育新种质的获得. 园艺学报 33, 374-377.
|
| 10 |
王娜, 王奎玲, 刘庆华, 刘庆超 (2017). 七种野生铁线莲属植物核型分析. 草业学报 26(11), 123-130.
|
| 11 |
王述彬, 罗向东, 戴亮芳, 钱春桃, 陈劲枫 (2004). 辣(甜)椒细胞质雄性不育系减数分裂和雄配子发生过程. 园艺学报 31, 807-810.
|
| 12 |
王文采, 李良千 (2005). 铁线莲属一新分类系统. 植物分类学报 43, 431-488.
|
| 13 |
王永周, 古松, 江莎, 任艳萍, 许珂, 李清河 (2008). 臭椿雄蕊的发育和雄配子体的形成. 园艺学报 35, 577-586.
|
| 14 |
王自芬 (2008). 毛茛科植物胚珠形态、结构、发育及其系统学意义. 博士论文. 西安: 陕西师范大学. pp. 25.
|
| 15 |
杨艺, 王娜, 马颖杰, 王奎玲, 刘庆华, 刘庆超 (2018). 4种铁线莲属植物种子萌发特性. 草业科学 35, 2395-2402.
|
| 16 |
杨艺, 王娜, 王奎玲, 刘庆华, 刘庆超 (2019). 铁线莲属4种植物的花芽分化研究. 园艺学报 46, 87-95.
|
| 17 |
张阿娟 (2006). 类叶升麻(毛茛科) 形态结构、发育及系统位置的研究. 硕士论文. 西安: 陕西师范大学. pp. 14-27.
|
| 18 |
张克中, 赵祥云, 梁励, 黄善武, 张启翔 (2003). 王百合雄性不育突变体小孢子败育的细胞学研究. 北京林业大学学报 25(3), 124-126.
|
| 19 |
张敏涛, 张荻, 申晓辉 (2016). ‘东方晨曲’铁线莲小孢子发生和雄配子体发育进程的解剖学研究. 西北植物学报 36, 2213-2219.
|
| 20 |
张燕, 黎斌, 李思锋 (2010). 铁线莲属植物分类学及园艺学研究进展. 中国野生植物资源 29(5), 6-10.
|
| 21 |
赵桦, 孟华菊, 徐娟 (2011). 七叶树小孢子发生及雄配子体发育研究. 西北植物学报 31, 217-222.
|
| 22 |
Bhandari NN (1968). Studies in the family Ranunculaceae. X. Embryology of Anemone L. Phytomorphology 18, 487-497.
|
| 23 |
Xie L, Li LQ (2012). Variation of pollen morphology, and its implications in the phylogeny of Clematis(Ranunculaceae). Plant Syst Evol 298, 1437-1453.
|
| 24 |
Zhang Q, Hao Q, Guo X, Liu QC, Sun YK, Liu QH, Wang KL (2017). Anther and ovule development in Camellia japonica(Naidong) in relation to winter dormancy: climatic evolution considerations. Flora 233, 127-139.
|

 )
)
 Home
Home