)
)
)
)
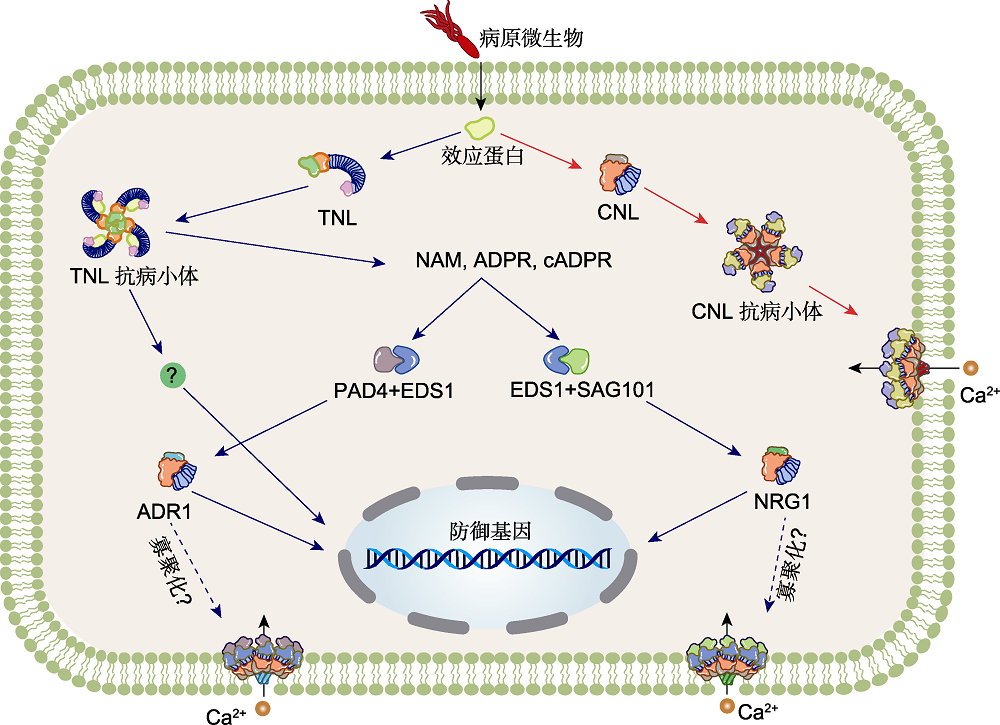
图3. 高等植物NLRs介导的免疫反应工作模型(改自Liu et al.,
当植物细胞被病原菌侵染时, 一些病原菌可分泌效应因子以突破植物的免疫防线。在长期进化过程中, 植物进化出许多胞内受体来识别这些效应因子, 从而促发其对病原菌的抗性。CNLs通过感知效应蛋白触发其在质膜的五聚化和抗病小体形成(该模型以间接识别模式的ZAR1抗病小体示例), 并通过N端CC结构域中的α1螺旋形成的孔道作为Ca2+内流通道, 介导胞质Ca2+浓度上升, 开启细胞死亡和防御反应。TNLs感知效应蛋白后形成四聚抗病小体(该模型以直接识别模式的RPP1示例), TNL抗病小体的形成导致TIR NADase激活, 触发可能包含EDS1-PAD4-ADR1s或EDS1-SAG101-NRG1s低聚合物的组装。辅助型NLR的寡聚作用可形成CNL类似孔道, 作为Ca2+内流通道, 介导下游免疫和细胞死亡。红色箭头表示CNL信号, 蓝色箭头表示TNL和RNL信号。
Figure 3. Working models of activation of NLRs-mediated immunity in higher plants (modified from Liu et al.,
Upon an infection of a plant cell by a pathogen, some pathogens can secrete effectors to break through the immune defense line of plants. In the process of long-term evolution, plants have evolved many intracellular receptors to recognize these effectors, so as to promote resistance to pathogens. CNLs triggers pentamerization and resistosome formation on the plasma membrane (PM) through sensing effector (The example here depicts a ZAR1 resistosome which indirect recognition effector assembly). The pore formed by the N-terminal CC α1 helices serves as a Ca2+ influx channel, mediating increase of cytosolic Ca2+ concentration and turning on cell death and defense responses. TNLs, upon perception of effectors (the model here depicts an example of direct effector-receptor recognition as with RPP1), formation of the TNL resistosome leads to activation of TIR NADase activity, triggering assembly of oligomeric complexes presumably containing EDS1-PAD4-ADR1s or EDS1-SAG101-NRG1s. The oligomerization of the helper NLRs enables a similar pore formation as CNLs, serving as Ca2+ influx channels to mediate downstream immunity and cell death.The red arrows indicate CNL signal, and the blue arrows indicate TNL and RNL signals.